



Введение
Изучение адаптивных особенностей волосяного покрова мелких лесных грызунов представляет большой научный интерес, и прежде всего с позиций эволюционной морфологии, экологической физиологии и таксономии этих животных. Освоение ими лесной подстилки и ходов неглубоких нор в условиях постоянного термического дефицита, а также общее свойственное им некомпенсируемое несовершенство терморегуляции способствовали выработке в процессе их эволюции целого комплекса экологических и морфофизиологических приспособлений, направленных на сохранение оптимального энергетического баланса организма со средой при низких и крайне неустойчивых окружающих температурах. Отсюда важность углубленных исследований таких адаптивных механизмов физической терморегуляции зверьков, как строение и теплозащитные свойства шерсти, микроструктура волос, сезонно-возрастная и популяционная изменчивость этих показателей, выявление особенностей строения шерстного покрова, связанных с обитанием в специфических температурных условиях приземного слоя воздуха, и т. п. Надо учитывать также и общую слабую изученность данного вопроса, особенно применительно к мелким лесным млекопитающим таежного Севера.
Материалы
Материалом для наших исследований послужили шкурки зверьков, отловленных в различные сезоны 1972–2014 гг. преимущественно в центральных и южных районах Карелии. Проанализировано 18 сухих коллекционных шкурок полевки-экономки и 26 – лесной мышовки, правда, с последней дело обстояло сложнее. В связи с тем что с сентября по вторую декаду мая она находится в зимней спячке (Ивантер, 1972, 1975, 2018), материалом для данной работы могли быть только шкурки зверьков, отловленных в летние месяцы. Среди них оказались экземпляры как с чистой мездрой и зрелым волосом, так и на различных стадиях линьки, с хорошо выраженной пигментацией кожи и подростом новых волос. Всего для изучения волосяного покрова рассматриваемого вида использовано 26 шкурок, которые и анализировали по стандартной методике. Кроме того, еще по 23 шкуркам лесной мышовки изучалась линька.
Методы
При исследовании волосяного покрова названных видов мелких млекопитающих использовались стандартные методы, неоднократно описанные в специальной литературе (Кузнецов, 1932, 1952; Церевитинов, 1951; Соколов, 1973; Соколов и др., 1988; Хмелевская, 1965), в т. ч. и в наших предыдущих публикациях (Ивантер, Левина, 1980, 1981, 1983; Ивантер и др., 1984, 1985; Ивантер, 2021). Цифровой материал обрабатывался статистически по общепринятой методике (Ивантер, 1979; Ивантер, Коросов, 1992, 2003; Коросов, 2007).
Результаты
Полевка-экономка – Microtus oeconomus Pall.
Общая характеристика волосяного покрова
Как и у других лесных грызунов, волосы у полевки-экономки растут пучками (по 4–7 в каждом) наклонно к поверхности кожи и образуют отчетливые потоки, обращенные к хвосту. Волосы лишены сегментации, укладываются в одном направлении и не свойлачиваются лишь благодаря многочисленности жестких и упругих направляющих волос, выполняющих в шерсти «разделительную» функцию. По сравнению с другими изученными видами мелких млекопитающих полевка-экономка обладает наиболее густым, длинным и пышным волосяным покровом. На спине – темно-бурый с легкой рыжиной и слабым черноватым налетом, образуемым темными окончаниями кроющих волос. Нередки экземпляры с большей примесью желтовато-ржавого оттенка или, наоборот, более темные с преобладанием черно-серых тонов. Бока черновато-бурые, по тону светлее спины, но гораздо темнее брюшка, окрашенного в тусклые серовато-белесые с желтоватым оттенком цвета. Из-за проглядывающих между белыми кончиками темных участков волос вся вентральная сторона тела выглядит грязно-белой или серой и резко контрастирует с темным верхом. У молодых зверьков волосы несколько короче, но гуще, чем у взрослых, и окрашены бледнее, в более серые и блеклые тона.
Дифференциация волос и их гистологическое строение
По М. Я. Марвину (1966), волосяной покров серых полевок Среднего Урала, в т. ч. и экономки, «состоит в основном из волос двух категорий: более длинных, толстых, но сравнительно редко расположенных и пигментированных волос – ости и более коротких, тонких, мягких, скрывающихся под остью, густо расположенных пуховых волос» (с. 45). Хотя автор и считает, что «остевые можно подразделить на направляющие и собственно ость, а пуховые – на промежуточные и собственно пуховые волосы», он от такого деления почему-то отказывается и цифровой материал приводит только по остевым и пуховым волосам. Иной точки зрения придерживается В. Е. Соколов (1973), выделяющий у полевки-экономки из окрестностей Иркутска волосы пяти типов (в пределах трех категорий): направляющие, остевые и пуховые, и те и другие двух порядков. Автор усматривает для каждого из них размерные и морфологические отличия. В то же время И. В. Назарова (1958) различает у серых полевок Волжско-Камского края четыре категории волос: направляющие, остевые, промежуточные и пуховые, а В. Г. Кривошеев с соавторами (1975), изучавший полевок-экономок северо-западной Чукотки, две: остевые и пуховые.
Результаты наших исследований в Карелии в общих чертах согласуются с данными В. Е. Соколова, с той лишь разницей, что волосы у наших полевок тоньше и чуть длиннее, кроме того, мы не делим остевые и пуховые на порядки, рассматривая лишь три категории волос: направляющие, остевые и пуховые (табл. 1, 2).
Таблица 1. Характеристика летней шерсти полевки-экономки
| Категория волос | n | Число волос на 4 кв. мм (М + m) |
Длина волоса, мм (М + m) | Толщина волоса, мкм (М + m) |
| Спина | ||||
| Направляющий | 9 | 10.3 ± 1.1 | 14.6 ± 0.04 | 54.0 ± 0.7 |
| Остевой | 14 | 22.6 ± 1.3 | 12.2 ± 0.05 | 49.1 ± 0.6 |
| Пуховой | 16 | 301.1 ± 3.0 | 10.9 ± 0.04 | 22.8 ± 0.7 |
| Бок | ||||
| Направляющий | 10 | 9.2 ± 1.2 | 13.3 ± 0.06 | 53.4 ± 0.7 |
| Остевой | 11 | 18.9 ± 1.8 | 11.9 ± 0.05 | 48.3 ± 0.8 |
| Пуховой | 15 | 259.9 ± 2.1 | 10.1 ± 0.03 | 21.7 ± 0.5 |
| Брюшко | ||||
| Направляющий | 9 | 10.1 ± 1.3 | 12.3 ± 0.08 | 51.4 ± 0.6 |
| Остевой | 9 | 21.4 ± 1.6 | 10.1 ± 0.06 | 47.1 ± 0.7 |
| Пуховой | 10 | 286.1 ± 2.2 | 8.6 ± 0.06 | 20.0 ± 0.6 |
Таблица 2. Характеристика зимней шерсти полевки-экономки
| Категория волос | n | Число волос на 4 кв. мм (М + m) |
Длина волоса, мм (М + m) | Толщина волоса, мкм (М + m) |
| Спина | ||||
| Направляющий | 10 | 18.4 ± 3.2 | 17.4 ± 0.07 | 52.7 ± 0.5 |
| Остевой | 9 | 26.0 ± 3.1 | 15.1 ± 0.05 | 48.6 ± 0.6 |
| Пуховой | 9 | 434.3 ± 5.1 | 12.3 ± 0.07 | 16.4 ± 0.3 |
| Бок | ||||
| Направляющий | 10 | 12.8 ± 4.1 | 15.8 ± 0.04 | 51.9 ± 0.6 |
| Остевой | 10 | 24.2 ± 3.6 | 14.7 ± 0.05 | 47.8 ± 0.05 |
| Пуховой | 10 | 415.6 ± 3.9 | 11.9 ± 0.05 | 16.0 ± 0.6 |
| Брюшко | ||||
| Направляющий | 10 | 14.1 ± 3.1 | 14.9 ± 0.06 | 50.1 ± 0.4 |
| Остевой | 16 | 26.9 ± 3.3 | 13.8 ± 0.06 | 45.9 ± 0.6 |
| Пуховой | 14 | 419.2 ± 3.6 | 11.0 ± 0.02 | 15.3 ± 0.6 |
Направляющие волосы возвышаются над основной массой шерсти и отличаются не только длиной, но также толщиной и формой стержня. В поперечнике они круглые, стержень прямой, без перетяжек, изгибов и продольных поворотов. От основания и примерно до середины волоса он сохраняет приблизительно одинаковую, относительно небольшую толщину, а в верхней части расширяется и постепенно переходит в хорошо выраженную веретеновидную гранну. Последняя сильно растянута по длине стержня, не имеет перетяжек и характеризуется длинным нитевидным кончиком, без сердцевины.
Кутикула направляющих волос некольцевидного типа и состоит из чешуек со слабо волнистым краем (рис. 1). В прикорневой зоне они сильно вытянуты, края их более зазубрены и относительно неплотно прилегают один к другому, а в гранне значительно расширяются, укорачиваются и теснее охватывают стержень. Строение сердцевины также меняется по длине стержня. У основания она начинается одним уплотненным рядом клеток, затем по мере расширения стержня постепенно становится четырехрядной и более рыхлой, с крупными внутри- и межклеточными полостями, заполненными воздухом и перемежающимися с зернами пигмента, после чего опять сужается до однорядной и полностью исчезает в концевой части. Клетки сердцевины на 3/4 длины волоса содержат пигмент черного цвета и лишь в конце гранны коричневого. В кончике волоса как сердцевина, так и пигментные зерна отсутствуют, поэтому он бесцветен.
Мощность коркового слоя направляющих волос достигает 9–11 мкм и обеспечивает им большую прочность в сравнении с волосами всех других категорий. В то же время по толщине сердцевины они не отличаются от остевых: этот слой занимает у них 86–89 % от общей толщины волоса (табл. 3).
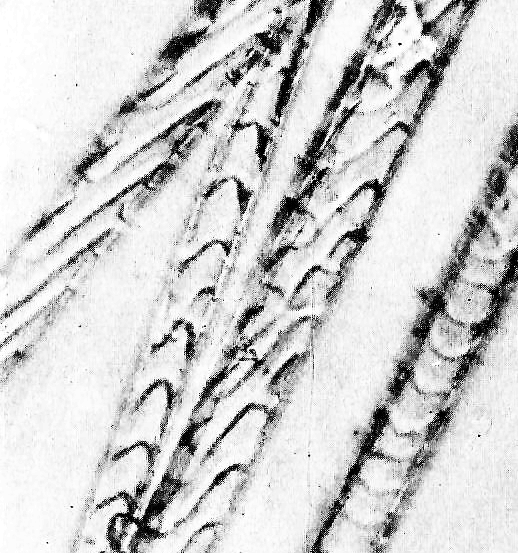
Рис. 1. Кутикула волоса полевки-экономки
Fig. 1. The cuticle of the root vole hair
Таблица 3. Толщина сердцевинного слоя стержня волос полевки-экономки
| Категория волос | Летняя шерсть | Зимняя шерсть | ||||
| n | толщина сердцевины, мкм (М + m) | % от общей толщины волоса | n | толщина сердцевины, мкм (М + m) | % от общей толщины волоса | |
| Спина | ||||||
| Направляющий | 9 | 46.6 ± 0.6 | 86.4 | 10 | 45.1 ± 0.4 | 85.6 |
| Остевой | 10 | 42.7 ± 0.5 | 87.0 | 9 | 42.7 ± 0.6 | 87.8 |
| Пуховой | 10 | 12.4 ± 0.7 | 54.3 | 9 | 13.4 ± 0.7 | 81.6 |
| Бок | ||||||
| Направляющий | 9 | 46.5 ± 0.4 | 87.0 | 9 | 44.7 ± 0.7 | 86.2 |
| Остевой | 11 | 41.8 ± 0.5 | 86.6 | 10 | 41.3 ± 0.5 | 86.4 |
| Пуховой | 10 | 12.0 ± 0.7 | 55.3 | 9 | 13.2 ± 0.6 | 82.8 |
| Брюшко | ||||||
| Направляющий | 9 | 45.0 ± 0.5 | 87.7 | 10 | 44.6 ± 0.4 | 89.0 |
| Остевой | 10 | 40.9 ± 0.7 | 86.9 | 16 | 40.6 ± 0.5 | 88.3 |
| Пуховой | 11 | 12.3 ± 0.6 | 61.1 | 14 | 12.9 ± 0.6 | 84.2 |
Количество направляющих волос у полевки-экономки невелико, однако гораздо больше, чем у землероек, а на спине и боках втрое меньше, чем у лесной мышовки. Летом их насчитывается от 8 до 12, зимой – от 13 до 21 на 4 мм2 шкурки, что от общего числа волос составляет 3.1–3.3 %. При этом от спины к брюшку плотность направляющих волос закономерно снижается. В том же порядке изменяется их длина и толщина: самые короткие и тонкие волосы растут на животе, а самые длинные и толстые – на дорсальной стороне тела (см. табл. 1, 2). Заметны и сезонные различия. В зимней шерсти направляющих не только больше, но они и длиннее, зато по толщине они несколько уступают им.
Остевые волосы значительно короче и тоньше направляющих и, кроме того, имеют перед гранной резкое сужение сердцевины, нередко с нарушением клеточной структуры и диффузным распределением пигментных зерен (так называемая шейка, или перетяжка). Гранна у них не такая длинная, как у направляющих, и занимает около 1/3 длины всего стержня. Форма ее ланцетовидная, уплощенная, кончик короткий, быстро утончающийся. В области шейки волос слегка поворачивается и изгибается, прикрывая расширенной гранной нижележащие пуховые волосы. По микроструктуре стержня остевые почти не отличаются от направляющих. Кутикула построена из чешуек некольцевидного типа (см. рис. 1). У основания волоса они расположены под острым углом друг к другу, сильно удлинены и имеют свободный зазубренный край, а в области гранны приобретают лентовидную форму и плотно охватывают стержень. Сердцевинный канал даже в самой широкой части гранны представлен лишь двумя рядами клеток. Расположены они очень рыхло и перемежаются крупными зернами пигмента и многочисленными воздушными полостями. У основания волоса и в области шейки сердцевина однорядная, а в самом кончике вообще не выражена. Корковый слой относительно тонкий и почти не меняется на протяжении всего стержня. Окраска остевого волоса зональная: от основания и до перехвата черного цвета, гранна – рыжего, а самый кончик – бесцветный.
Количество остевых волос у экономок в 2–3 раза больше, чем направляющих: на 4 мм2 шкурки их насчитывается 16–32 летом и 22–60 зимой (5.4–6.8 % от общего числа волос всех категорий). Меняется по сезонам и строение остевых. Зимой они на 2.8–3.7 мм длиннее и на 0.5–1.2 мкм тоньше, чем летом. При этом и в зимней, и в летней шерсти наиболее длинные, толстые и густые остевые волосы растут на спине, а самые короткие, тонкие и редкие – на брюшке.
Пуховые волосы самые тонкие, короткие и извитые, со слабо выраженной гранной. Толщина их одинакова на протяжении всего стержня, но в последнем изгибе имеется сужение сердцевины без продольного поворота. В этом месте располагается цветовая граница волоса: от основания до перехвата он черный, а от перехвата до кончика или рыжий (на спине и боках), или белый (на брюшке). Рисунок кутикулы приблизительно такой же, как у остевых и направляющих, но построена она из более мелких чешуйчатых клеток и имеет почти одинаковое строение по всей длине волоса. Корковый слой у пуховых выражен слабо, этим, очевидно, и объясняется относительная непрочность волоса на разрыв. Сердцевина у основания волоса фрагментарная, иногда совсем не выражена, а затем появляется в виде расположенного лестницей одного ряда клеток, в таком относительно неизменном виде присутствует на всем протяжении волоса. В летних волосах на долю сердцевины приходится 54–61 % ширины стержня, а в зимних – 82–84 %. Утолщение мозгового слоя волоса происходит в основном за счет воздушных полостей, что вместе с увеличением густоты и длины пуховых волос способствует накоплению в подпуши (между волосками и в самих волосах) воздушной прослойки и определяет лучшие теплозащитные свойства зимней шерсти.
Обращает внимание удивительное постоянство доли, которую занимают пуховые волосы в летнем и зимнем волосяном покрове полевки-экономки. Летом на спине, боках и брюшке она составила соответственно 90.21, 90.3 и 90.0 % от общего числа волос, а зимой – 90.6, 91.6 и 91.0 %. В то же время в абсолютных показателях численность пуховых значительно возрастает от лета к зиме и снижается по направлению от спины к брюшку (см. табл. 1, 2). Приблизительно с такой же закономерностью изменяется длина и толщина пуховых волос. Летом они значительно короче и тоньше, чем зимой, и во все сезоны уменьшаются в длину и толщину в дорсовентральном направлении.
Плотность волосяного покрова
Волосы у полевки-экономки распределяются по телу относительно равномерно. Однако на спине они несколько гуще, чем на брюшке и боках (см. табл. 1, 2). При этом, если, по М. Я. Марвину (1966, 1974), наибольшей изреженностью волос отличается брюшко, то, по нашим данным, это свойственно бокам. Сезонная изменчивость плотности волос происходит преимущественно за счет пуховых. Среди зверьков, отловленных осенью, попадаются особи с так называемым двухъярусным волосяным покровом: на фоне старых волос хорошо заметны новые, достигающие более половины длины старых. Плотность волос зимой возрастает в 1.4 раза по сравнению с летом. Между тем осенний наряд зверьков, состоящий из невыпавших старых волос и подрастающих новых, плотнее зимнего. Летом общее число волос (на 4 мм2) на спине составляет 334, на боках – 288, на брюшке – 318. Зимой соответственно 479, 453 и 460. Тем не менее соотношение кроющих (направляющих и остевых) и пуховых волос во все сезоны остается примерно одинаковым: 1:9 – 1:10.
Обращает внимание относительно более густая опушенность брюшка у экономок в сравнении с боками, что обычно характерно для видов, ведущих полуводный образ жизни (Церевитинов, 1958), и было зафиксировано нами у водяной куторы (Ивантер и др., 1984). Строго говоря, полевка-экономка к этой категории не относится, однако принадлежит к наиболее стенотопным видам грызунов и предпочитает увлажненные биотопы, приуроченные к травяным болотам и водоемам. Она хорошо плавает, не боится воды и по гидрофильности близка к водяной полевке (Ивантер, 1975). Этим, вероятно, и объясняется описанное выше необычное расположение волос на теле зверька.
Толщина волос
Как и у большинства других мелких млекопитающих, толщина волос у экономки варьирует в зависимости от местоположения на шкурке и различна у волос разных типов. Самые толстые волосы направляющие, средняя толщина у остевых, а наиболее тонкие – пуховые (см. табл. 1, 2). На спине волосы всех категорий как зимой, так и летом достигают максимальной толщины, а на брюшке – минимальной. Изменяется толщина волос и по сезонам. При этом у направляющих и остевых диаметр стержня уменьшается от лета к зиме всего на 0.5–1.5 мкм (1–3 %), а у пуховых – на 4.9–6.4 мкм (24.3–28.0 %), т. е. гораздо существенно. Что же касается сердцевины волос, то у остевых и направляющих относительная толщина ее к общему диаметру волоса и зимой, и летом остается приблизительно одинаковой, тогда как у зимних пуховых этот показатель на 23–27 % больше, чем у летних (см. табл. 3). Корковый слой у волос всех категорий развит у этого вида слабо, особенно в сравнении с землеройками. Так, если у кроющих волос обыкновенной бурозубки он занимает вместе с кутикулой от 29 до 50 % общей толщины стержня (Ивантер, 2021), то у полевки-экономки – всего 6–7 %.
Длина волос
По высоте волосяного покрова на шкурке экономки выделяется четыре зоны: крестец, где растут наиболее длинные волосы, большая часть спины со средней длиной волос около 15 мм, бока, характеризующиеся умеренной высотой волос, и, наконец, брюшко, на котором находятся самые короткие волосы (рис. 2). Согласно номенклатуре Б. Ф. Церевитинова (1958), такой тип топографии по высоте волос называется сакральным и относится к числу наиболее распространенных у наземных млекопитающих. Хорошо выражены сезонные изменения рассматриваемого показателя: зимняя шерсть экономок по всем категориям волос на 2–3 мм длиннее летней (см. табл. 1, 2). У направляющих волос этот прирост составляет на разных участках шкурки 21.5–2.8 мм (19–21 % от летних показателей), у остевых – 2.8–3.7 мм (23.5–36.6 %), у пуховых – 1.4–2.4 мм (12.8–28.0 %). Из всех изученных нами видов мелких млекопитающих полевка-экономка отличается наиболее высоким волосяным покровом. Так, на спине длина остевых волос летом достигает у нее в среднем 12.2 мм, зимой – 15.1 мм, это почти на 5 мм больше, чем у превышающего ее по весу и размерам тела обыкновенного крота (Ивантер, Моисеева, 2022).
Замечена и географическая изменчивость данного показателя (табл. 4): у полевок-экономок из окрестностей Иркутска длина остевых волос на спине достигает летом 12.5 мм, зимой – 15.0, в Карелии соответственно 12.2 и 15.1, в Чаунской тундре Чукотки – 13.1 и 17.0, на Среднем Урале летом – 15.3 мм. В то же время длина пуховых волос варьирует по регионам в более узких пределах. При этом идентичность примененных разными авторами методик измерений волос позволяет отнести выявленные различия за счет популяционных.
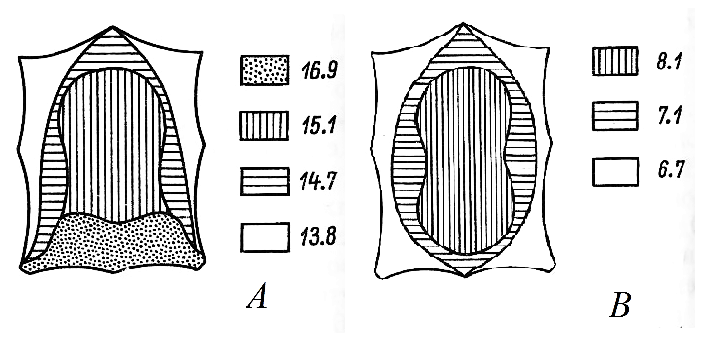
Рис. 2. Топография высоты волос полевки-экономки (А) и лесной мышовки (В)
Fig. 2. Topography of the hair height of the root vole (A) and the north birch mouse (B)
Таблица 4. Географические различия летнего волосяного покрова полевки-экономки
(по средним данным)
| Место исследования | Число волос на 4 кв. мм |
Относительное количество, пуховых, % | Длина волос, мм | Толщина волос, мкм | ||||||
| направляющий | остевой | пуховой | направляющий | остевой | пуховой | Литературный источник | ||||
| Карелия | 313.2 | 90.2 | 13.4 | 11.4 | 9.9 | 52.9 | 48.2 | 21.6 | Наши данные | |
| Волжско-Камский край | 338.4 | 88.7 | 17.9* | 13.3* | 11.0* | – | – | – | Назарова, 1958 | |
| Средний Урал | 266.3 | 95.3 | 15.3 | 11.4 | 54.9 | 10.7 | Марвин, 1974 | |||
| Окрестности г. Иркутска | – | – | 14.3 | 12.5 | 10.1 | 49.0 | 50.0 | 21.0 | Соколов, 1973 | |
| Северо-Западная Чукотка | 259.8 | 81.8 | 13.1 | 9.4 | – | – | – | Кривошеев и др., 1975 | ||
Примечание. * – максимальные размеры волос.
Теплоизоляционные свойства покровов
Разобранные выше структурные особенности волосяного покрова полевок во многом определяют его термозащитные свойства. Относительно большая густота, длина, толщина волос и развитие сердцевинного слоя способствуют снижению теплопроводности шкурки и, следовательно, улучшению ее термоизоляционных способностей. Это особенно важно для полевки-экономки – вида, обитающего в сырых, захламленных, плохо прогреваемых солнцем местах и испытывающего здесь определенные трудности в поддержании теплового баланса. Вместе с тем в развитии густого и относительно толстого волосяного покрова полевки-экономки проявляется одна из сторон высокой приспособленности этого вида к существованию в районах с холодным климатом и относительное его распространение в Субарктике (Кривошеев и др., 1975).
Как и у прочих мелких млекопитающих, зимняя шкурка полевки-экономки обладает более высокими термоизоляционными свойствами, чем летняя (табл. 5). Коэффициент теплопроводности зимней шкурки уменьшается по сравнению с летней на 11.3 единицы измерения, что составляет 23.6 %. Напомним, что густота и длина волос увеличивается у полевок к зиме примерно на ту же величину (13–30 %). Тем не менее повышенными теплозащитными качествами отличаются и шкурки в период линьки. Как и в отношении других видов землероек и грызунов, мы связываем это с утолщением кожи во время линьки и увеличением густоты меха за счет подроста новых волос при частичном сохранении старых.
Таблица 5. Коэффициент теплопроводности (10–3 Вт/м К) сухих шкурок лесной мышовки и полевки-экономки
| Сезон | n | Пределы | M ± m |
| Лесная мышовка | |||
| Лето | 22 | 48.0–50.3 | 48.6 ± 0.3 |
| Осень | 4 | 37.9–38.2 | 38.1 ± 0.4 |
| Полевка-экономка | |||
| Лето | 11 | 46.7–53.2 | 47.8 ± 0.4 |
| Осень | 8 | 34.1–40.9 | 36.1 ± 0.6 |
| Зима | 5 | 34.1–38.4 | 36.5 ± 0.4 |
На основании детальных исследований структуры волосяного покрова, его удельной теплопроводности и микроструктуры стержня волоса, выполненных на исследованных шкурках лесной мышовки, выявлены приспособительные особенности ее шерсти, связанные со своеобразием видовой экологии и, в частности, с наличием зимней спячки. Обитая преимущественно в лесной подстилке, среди упругих стеблей травы и в узких ходах неглубоких нор, но появляясь нередко и на поверхности почвы, эти зверьки приобрели в процессе эволюции более редкий и грубый, но длиннее и менее выровненный, чем, например, у землероек и полевок, шерстный покров, характеризующийся отчетливыми потоками волос в направлении от головы к хвосту и заметно выраженной трехслойностью. Более редкий поверхностный слой образуют относительно толстые и длинные направляющие волосы, средний построен из зонально окрашенных остевых, а нижний – из тонких и волнистых пуховых. Волосяной стержень, не образуя сужений сердцевины, утолщается от основания к хорошо выраженной гранне, а затем переходит в тонкий нитевидный кончик. Форма стержня веретеновидная, без изгибов, продольных поворотов и перетяжек. Кутикула некольцевидного типа и представлена чешуйчатыми клетками с волнистым краем, а корковый слой не меняется по толщине почти на всем протяжении стержня и состоит из расположенных рядами чечевицеобразных клеток сердцевины. Особи в состоянии линьки встречаются в течение всего лета. У взрослых, перезимовавших, она проходит в июне – июле, а у прибылых – в августе – сентябре.
Линька
Согласно нашим исследованиям, полевка-экономка на протяжении жизни претерпевает одну возрастную (ювенильную) и две сезонные (весеннюю и осеннюю) линьки (рис. 3).
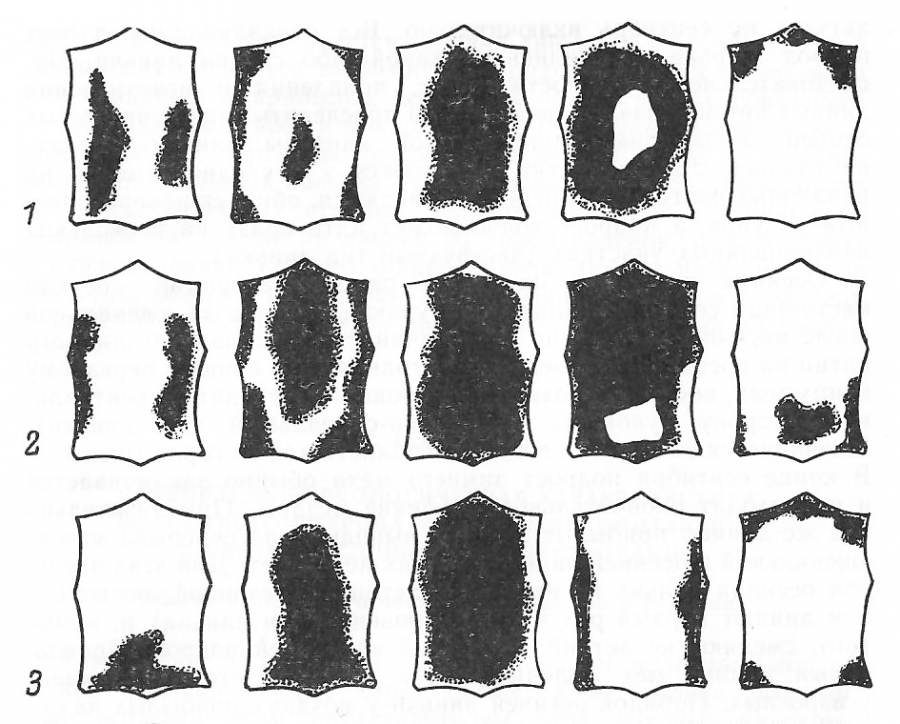
Рис. 3. Последовательность появления линных пятен у полевки-экономки: 1 – ювенильная линька; 2 – весенняя; 3 – осенняя
Fig. 3. The sequence of the appearance of moulting spots in the root vole: 1 – juvenile molt; 2 – spring moult; 3 – autumn moult
Ювенильная линька характеризуется полной сменой детского волосяного покрова на взрослый. В зависимости от того, в какое время года она протекает (а это, в свою очередь, зависит от сроков появления на свет прибылых зверьков), ее можно считать весенней, летней или осенней, а шерстный покров, формирующийся в процессе этой линьки, – соответственно летним, осенним или зимним, т. к. он по своим морфометрическим показателям ничем не отличается от наряда взрослых. Происходит ювенильная линька после окончания подрастания «детского» меха и перехода зверьков к самостоятельной жизни по выходе из гнезда. В связи с растянутостью размножения полевок-экономок с конца апреля по сентябрь (Ивантер, 1975, 2018) молодых на разных стадиях ювенильной линьки можно встретить в течение всего лета. Подрост новых волос при этой линьке соответствует так называемому сублатеральному типу (Крыльцов, 1962). Начинается она обычно на боках тела с постепенным переходом на дорсальную и вентральную стороны, однако раньше других успевают сформироваться волосы брюшка (вероятно, потому, что здесь они короче и на их рост требуется меньше времени, чем на других участках), затем боков и в последнюю очередь задней части спины. Судя по состоянию мездры, завершается процесс на крестце или на затылке (см. рис. 2).
Осенняя линька прибылых и перезимовавших полевок в наших условиях наиболее интенсивно происходит с третьей декады августа по сентябрь включительно. Все отлавливаемые в этот период зверьки находились на какой-либо стадии линьки. Последовательность подроста волос, появления и исчезновения линных пятен оказалось возможным проследить лишь у прибылых особей. У перезимовавших четкой картины линьки выявить не удалось. Линные пятна появляются одновременно на различных частях тела, нередко сливаются, образуя неопределенный рисунок, а подрост волос может идти сразу на нескольких изолированных участках (диффузный тип линьки).
Осенняя линька у прибылых ранних генераций – первая настоящая сезонная линька, наступающая вслед за ювенильной после месячного перерыва. Она начинается с проявления линного пятна на крестце, которое, распространяясь на спину к переднему концу тела, постепенно охватывая бока, переходит на вентральную сторону туловища (цефально-сакральный тип линьки). Завершается смена волос в паховой области или на горле (см. рис. 2). В конце сентября подрост зимнего шерстного покрова обычно заканчивается, и происходит окончательное очищение мездры. Приблизительно так же линяют прибылые поздних выводков, но перерыва между ювенильной и осенней линьками у них не бывает. Для этих зверьков осенняя линька не является настоящей сезонной, поскольку они линяют первый раз в жизни (ювенильная линька) и, кроме того, сменяют не летний, а детский волосяной покров. Правда, новый зимний наряд молодых ничем не отличается от зимнего у взрослых. Порядок осенней линьки у поздних прибылых не соответствует последовательности смены покровов, описанной для выводков, а повторяет ход ювенильной линьки, наблюдавшейся у этих последних летом (сублатеральный тип линьки).
Весенняя линька полевки-экономки происходит в последовательности, обратной осенней, т. е. подрост новых и выпадение старых волос начинается в первую очередь на брюшке, хотя линное пятно и появляется сначала на боках. Дело в том, что волосы брюшка в наиболее широкой их части белые, поэтому в начальный период линьки в коже брюшка отсутствует пигмент, несмотря на то, что она утолщена и в ней происходят соответствующие морфообразовательные процессы. В противоположность этому на боках и спине кожа темнеет сразу, т. к. пигмент концентрируется в ней одновременно с возобновлением функционирования волосяных фолликулов.
Популяция полевок-экономок в период весенней линьки бывает, как правило, неоднородной в возрастном отношении, чем и объясняется растянутость этого процесса с апреля по вторую декаду июля. Первыми вступают в линьку зимовавшие зверьки, затем прибылые ранних выводков (после ювенильной линьки). У зимовавших она продолжается до конца июня и в июле в основном заканчивается. Однако особи с небольшими по площади темными пятнами на мездре, обычно на затылке и боках, встречаются нередко и в августе. У сеголеток ранних выводков смена ювенильного наряда на взрослый заканчивается в середине июля, а в августе начинается осенняя линька, в процессе которой летний волос сменяется на зимний. Порядок весенней смены покровов у зимовавших и прибылых полевок в общих чертах совпадает.
С. И. Огнев (1950) и К. Бауэр (Bauer, 1953) указывают на сходство в общих чертах возрастной и сезонной линек у полевки-экономки и водяной полевки. А. И. Крыльцов (1958) усматривает это сходство еще у стадных полевок и степной пеструшки и, опираясь на данные по линьке 20 видов полевок, делает вывод о существовании единого общего плана линьки полевок родов Microtus, Lagurus и Alticola. От себя добавим, что по характеру и последовательности линьки полевка-экономка сходна и с рыжей полевкой.
На основании детальных исследований структуры волосяного покрова, его удельной теплопроводности и микроструктуры стержня волоса, выполненных на 26 шкурках лесной мышовки, выявлены приспособительные особенности ее шерсти, связанные со своеобразием видовой экологии и, в частности, с наличием зимней спячки. Обитая преимущественно в лесной подстилке, среди упругих стеблей травы и в узких ходах неглубоких нор, но появляясь нередко и на поверхности почвы, эти зверьки приобрели в процессе эволюции более редкий и грубый, но длиннее и менее выровненный, чем, например, у землероек и полевок, шерстный покров, характеризующийся отчетливыми потоками волос в направлении от головы к хвосту и заметно выраженной трехслойностью. Более редкий поверхностный слой образуют относительно толстые и длинные направляющие волосы, средний построен из зонально окрашенных остевых, а нижний – из тонких и волнистых пуховых. Волосяной стержень, не образуя сужений сердцевины, утолщается от основания к хорошо выраженной гранне, а затем переходит в тонкий нитевидный кончик. Форма стержня веретеновидная, без изгибов, продольных поворотов и перетяжек. Кутикула некольцевидного типа и представлена чешуйчатыми клетками с волнистым краем, а корковый слой не меняется по толщине почти на всем протяжении стержня и состоит из расположенных рядами чечевицеобразных клеток сердцевины. Особи в состоянии линьки встречаются в течение всего лета. У взрослых, перезимовавших, она проходит в июне – июле, у прибылых – в августе – сентябре.
Лесная мышовка – Sicista betulina Pall.
Общая характеристика волосяного покрова
В связи с тем, что лесная мышовка на протяжении всей исследованной нами территории с сентября по вторую декаду мая находится в зимней спячке (Ивантер, 1972, 1975, 2018), материалом для данной работы могли быть только шкурки зверьков, отловленных в летние месяцы. Среди них оказались экземпляры как с чистой мездрой и зрелым волосом, так и на различных стадиях линьки, с хорошо выраженной пигментацией кожи и подростом новых волос.
Лесная мышовка (Sicista betulina) – единственная из всех мелких лесных млекопитающих таежного Севера относится к семейству тушканчиковых и является зимоспящим видом. Это обстоятельство определяет особый интерес к изучению механизмов ее физической терморегуляции, тем более что соответствующих исследований по данному вопросу практически не проводилось.
Общая характеристика шерстного покрова
Среди всех исследуемых нами грызунов лесная мышовка выделяется яркой рыже-желтой окраской с узкой полосой, идущей по линии хребта. Общий тон верха варьирует от бледного серо-палевого до охристо-рыжеватого, бока ржаво-желтоватого цвета, брюшко желтовато-серое или ржаво-белесое. Основной рыжеватый фон окраски слегка подернут черноватой вуалью, образуемой терминальными частями зонально окрашенных кроющих волос. Основания их аспидно-серые, средние части ярко-рыжеватого или охристого оттенка, а самые кончики черно-бурые или (у самых длинных волос) почти совсем черные. Волосы, образующие подпушь, однотонного серого цвета, более темные в верхней части и бледнеющие к основанию.
Волосяной покров у лесной мышовки более редкий и грубый, но длиннее, чем у землероек. Кроме того, он менее выровнен и характеризуется отчетливыми потоками полос в направлении от головы к хвосту. В волосяном покрове отчетливо выделяется три слоя – более редкий поверхностный, образуемый толстыми и длинными направляющими волосами, средний, построенный из зонально окрашенных остевых, и нижний – из тонких и волнистых пуховых. Стержни волос имеют слегка извитое основание. Они растут наклонно к поверхности кожи, но пучков и групп, характерных для большинства представителей отряда грызунов, в т. ч. и степной мышовки (Соколов, 1973), не образуют, т. е. расположение волос одиночное.
В волосяном покрове мышовок можно выделить три категории волос: направляющие, остевые и пуховые (рис. 4). Направляющие волосы наиболее длинные, упругие и толстые (табл. 6). Стержень, не образуя сужений сердцевины, постепенно утолщается от основания к хорошо выраженной гранне, а затем переходит в тонкий нитевидный кончик. Форма стержня веретеновидная, без изгибов, продольных поворотов и перетяжек. Гранна не растянута вдоль всего стержня, а занимает примерно четверть его длины.
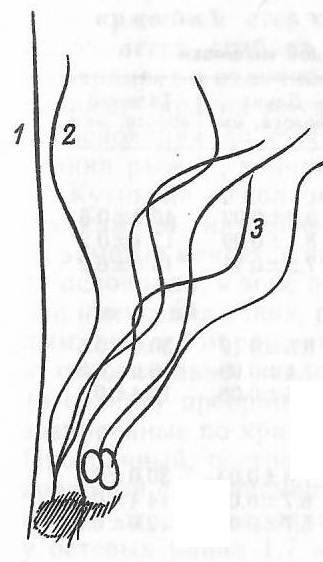
Рис. 4. Летние волосы лесной мышовки: 1 – направляющий, 2 – осевой, 3 – пуховой
Fig. 4. Summer hair of the northern birch mouse: 1 – guide, 2 – axial, 3 – down
Таблица 6. Характеристика летней шерсти лесной мышовки
| Категория волос | n | Число волос на 4 кв. мм (М + m) |
Длина волоса, мм (М + m) | Толщина волоса, мкм (М + m) |
| Спина | ||||
| Направляющий | 26 | 36.1 ± 1.4 | 9.0 ± 0.02 | 40.0 ± 0.6 |
| Остевой | 25 | 97.2 ± 4.8 | 8.1 ± 0.09 | 17.4 ± 0.2 |
| Пуховой | 27 | 443.0 ± 6.1 | 7.5 ± 0.11 | 15.8 ± 0.2 |
| Бок | ||||
| Направляющий | 25 | 24.8 ± 1.3 | 8.5 ± 0.12 | 30.0 ± 0.5 |
| Остевой | 25 | 60.6 ± 4.6 | 7.1 ± 0.09 | 16.0 ± 0.5 |
| Пуховой | 25 | 410.3 ± 5.2 | 6.5 ± 0.08 | 15.4 ± 0.4 |
| Брюшко | ||||
| Направляющий | 25 | 10.9 ± 1.8 | 7.8 ± 0.03 | 30.0 ± 0.5 |
| Остевой | 25 | 47.3 ± 4.1 | 6.7 ± 0.04 | 14.4 ± 0.03 |
| Пуховой | 24 | 323.7 ± 2.9 | 5.7 ± 0.04 | 12.0 ± 0.3 |
Дифференциация волос и их гистологическое строение
Кутикулярный слой представлен чешуйчатыми клетками некольцевидного типа, вытянутыми, остроконечными у основания волоса и значительно укороченными и уплотненными в самой широкой его части. Корковый слой, толщина которого не меняется почти на всем протяжении стержня и составляет около 4 мкм, обеспечивает упругость и прочность волоса и служит каналом для расположенных рядами чечевицеобразных клеток сердцевины. Ширина сердцевинного канала, начинающегося у основания волоса одним рядом клеток, достигает максимума в середине гранны (31.8 мкм), где клетки сердцевины расположены в 3–4 ряда, в кончике волоса сердцевина отсутствует.
Большая часть гранны направляющего волоса спины содержит темно-коричневый пигмент, а на боках и брюшке – светло-коричневый, почти рыжий. Ножка (основание) стержня на любом участке шкурки всегда черная, темнее остальных частей окрашены обычно и самые кончики волос.
Количество направляющих волос в сравнении с другими невелико (3–7 % от общего числа волос), они распределены на туловище неравномерно. Больше всего их на спине (от 14 до 40 на 4 мм2), меньше – на боках (10–34) и совсем немного – на брюшке (4–12). Так же происходит и укорочение направляющих волос: на спине длина их достигает в среднем 9.0 мм, на боку – 8.5, на брюшке – 7.8 мм.
Остевые волосы, в отличие от одноименных волос насекомоядных, не сегментированы. По сравнению с направляющими они характеризуются хорошо выраженной, рельефной гранной и заметным сужением стержня перед ней. В месте сужения (шейке) волос слегка загибается, но продольного поворота не образует и сохраняет в отличие от насекомоядных клеточное строение сердцевины. Окраска остевых волос, как и направляющих, зональная: от основания до сужения аспидно-серая, иногда почти черная, гранна рыжая, кончики волос бесцветные.
Кутикула некольцевидного типа представлена чешуйчатыми клетками вытянутой формы с волнистым краем (рис. 5). Строение их заметно меняется на протяжении волоса. В тонких его участках (у основания, в зоне перехода в гранну и в терминальных частях) они имеют вид узких, ромбических пластинок со свободным округлым краем, обращенным к вершине волоса, сильно удлинены и относительно неплотно охватывают стержень, а в широких (в гранне) превращаются в очень короткие и широкие чешуйки, зазубренные по краям и более плотно прилегающие одна к другой.

Рис. 5. Микроструктура волос лесной мышовки (ув. х600): а – кутикула, б – стержень волоса
Fig. 5. Microstructure of the hair of the northern birch mouse (x600): a – cuticle, б – hair shaft
Прозрачный, построенный из удлиненных веретеновидных клеток корковый слой пигмента не содержит и развит в сравнении с корковым слоем направляющих волос гораздо слабее. Толщина его у остевых волос равна 1.7 мкм, т. е. в 2 с лишним раза меньше, чем у направляющих. Сердцевинный канал просматривается на протяжении всего стержня, кроме кончика, в котором он вообще отсутствует, и прикорневой части, где сердцевина или не выражена (зрелый волос), или очень узкая, однорядная, почти незаметная даже под микроскопом (растущий волос). Чуть выше основания сердцевина становится отчетливо двухрядной, а затем число рядов увеличивается и достигает 4–5 в гранне (см. рис. 5). Параллельно этому возрастает и диаметр сердцевинного канала, достигающий в наиболее расширенной части 79–80 % от общей толщины волоса (см. табл. 2). Цвет зависит от концентрации пигментных зерен, расположенных в клетках сердцевины, и от распределения пигмента как по длине волоса, так и на разных частях тела.
Количество остевых волос на разных участках шкурки неодинаково и меняется в том же порядке, что и направляющих: больше всего их на спине (38–121 на 4 мм2), далее следуют бока (36–81) и брюшко (31–60). В этом же направлении изменяется и длина волос: на спине – 8.1 мм, на боках – 7.1, на брюшке – 6.7 мм.
Пуховые волосы, составляющие наиболее многочисленную группу (см. табл. 6), отличаются слабо развитой гранной, можно даже сказать, что она совсем отсутствует. Тонкий и относительно короткий стержень образует четыре сужения, самое заметное из которых находится непосредственно перед гранной (т. е. в верхней части волоса). Сердцевина, как правило, однорядная по всей длине стержня, и хотя в местах сужений она заметно утончается, клеточная структура в ней сохраняется. Окраска волос этой категории, равно как и других, двуцветная: рыжая в гранне и черная от основания волоса до последнего сужения стержня. Волос в целом выглядит слегка извитым, однако четкой сегментации, как у землероек, не имеет. В частности, отсутствуют продольные повороты стержня и прерывистое строение сердцевинного канала. Что же касается микроструктуры пуховых волос, то она ничем существенным не отличается от описанной для направляющих и остевых. Тем не менее можно отметить относительно слабое развитие коркового слоя. Средняя толщина его у пуховых волос составляет 1.5 мкм, таким образом, он в 1.2 раза тоньше, чем у остевых.
Выявляются особенности и в строении сердцевинного канала (табл. 7). В отличие от волос других категорий сердцевина пуховых более развита и построена из одного ряда рыхло соединенных и перемежающихся воздухоносными полостями пигментированных клеток. Относительно большая толщина сердцевины и насыщенность ее инертным воздухом определяют значение пуховых волос в осуществлении терморегуляции и защите организма от потери тепла. Волосы других категорий существенно уступают им в этом отношении, так что теплозащитные свойства волосяного покрова лесной мышовки зависят в основном от слоя пуховых волос. Последнее связано и с их количеством: на разных участках шкурки пуховые волосы составляют 82–85 % от волос всех категорий. При этом густота, длина и толщина пуховых волос зависят от их местоположения на туловище грызуна и меняются согласованно: самые густые, толстые и длинные волосы растут на спине, а самые редкие, тонкие и короткие – на брюшке. Пуховые же на боках тела занимают по этим показателям промежуточное положение.
Таблица 7. Толщина сердцевинного слоя стержня волос лесной мышовки
| Категория волос | n | Толщина сердцевины, мкм (М + m) | % от общей толщины волоса |
| Спина | |||
| Направляющий | 16 | 31.8 ± 0.4 | 79.7 |
| Остевой | 20 | 13.9 ± 0.4 | 79.9 |
| Пуховой | 18 | 12.8 ± 0.3 | 81.0 |
| Бок | |||
| Направляющий | 15 | 23.4 ± 0.3 | 77.9 |
| Остевой | 17 | 12.6 ± 0.5 | 78.8 |
| Пуховой | 18 | 12.1 ± 0.4 | 79.1 |
| Брюшко | |||
| Направляющий | 16 | 22.8 ± 0.4 | 76.0 |
| Остевой | 16 | 11.4 ± 0.3 | 78.9 |
| Пуховой | 16 | 11.3 ± 0.3 | 77.8 |
Плотность волосяного покрова
При кажущейся равномерности распределения волос на теле зверька наблюдается постепенное изреживание шерстного покрова в направлении от спины к брюшку (см. табл. 6). Общее число всех волос (на 4 мм2) на спине составило в среднем 576 + 8, на боку – 496 + 3, на брюшке – 382 + 7, причем такой же порядок изменения густоты волос от спины к брюшку наблюдается и у волос всех категорий в отдельности. Низкая плотность волос на брюшке, по всей видимости, не сказывается отрицательно на теплопроводности шерстного покрова в целом, т. к. периоды похолодания зверьки этого вида обычно проводят в состоянии оцепенения, принимая характерную позу спячки: свернувшись клубочком, уткнув мордочку и лапки в живот и обвязавшись хвостом так, что открытой остается только спинная сторона тела, опушенная более густыми и длинными волосами.
Как и у большинства мелких млекопитающих, основную массу волос мышовки составляют пуховые (на спине – 76.1 %, на боках – 82.7, на брюшке – 85.0 %), на долю остевых приходится 10–12 %, а направляющие настолько малочисленны (3–6 % от общего числа волос всех категорий), что в формировании волосяного покрова существенной роли не играют. Им, очевидно, принадлежит осязательная и отчасти укрепляющая шерстный покров функция (Кузнецов, 1932, 1952; Церевитинов, 1951).
Толщина волос
Уменьшается толщина волос всех категорий в том же дорсовентральном направлении, что и их густота (см. табл. 6). Наиболее толстые волосы располагаются на спине, наиболее тонкие – на брюшке. Сердцевина же развита у них примерно одинаково и составляет от 76 до 81 % общей толщины стержня.
Наибольшей толщиной отличаются направляющие волосы, за ними в порядке убывания следуют остевые и пуховые, причем особенно велико отличие направляющих от двух других (оно достигает 2.5-кратной величины), тогда как эти последние различаются между собой незначительно. Тем не менее и здесь сердцевина характеризуется близкими относительными показателями (см. табл. 6).
Длина волос
Волосяной покров лесной мышовки по сравнению с другими видами исследованных грызунов (полевками, хомяками, мышами и др.) невысокий: на спине длина остевых волос составляет в среднем 8.1 мм, на боках – 7.1, на брюшке – 6.7 мм. В то же время у рыжих полевок эти показатели выше приблизительно на 10–12 %, а у серых – на 50–67 %. Что же касается длины волос других категорий, то они по данному признаку выстраиваются в тот же ряд, что и по толщине: самые длинные – направляющие (7.2–9.9 мм), а самые короткие – пуховые (5.1–7.7 мм). Остевые занимают в этом ряду промежуточное положение. При этом обращает внимание заметное укорочение волос всех категорий в дорсовентральном направлении (см. табл. 6).
Таким образом, по высоте и густоте волос на шкурке лесной мышовки отчетливо выделяются три зоны (см. рис. 2). Наиболее развит волосяной покров на спине, а по направлению к голове, брюшку и крестцу длина и густота волос снижаются. Согласно номенклатуре Б. Ф. Церевитинова (1958), такой тип распределения волос на шкурке называется дорсальным и встречается у горностаев, хомяков и тушканчиков. Напомним, кстати, что лесная мышовка единственная из наших лесных мышевидных грызунов относится к семейству тушканчиковых, от которых она унаследовала не только зимнюю спячку, но и своеобразную топографию шерстного покрова.
Теплоизоляционные свойства покровов
Измерение коэффициента удельной теплопроводности сухих шкурок исследованных мелких млекопитающих показало (см. табл. 5), что лучшими термозащитными качествами обладают шкурки в разгар осенней линьки. Коэффициент теплопроводности их оказался в 1.3 раза ниже, чем шкурок с уже сформированным волосяным покровом без следов линьки. Не оказалась в этом отношении исключением и лесная мышовка. У нее, как и у других исследованных видов, это можно объяснить значительным утолщением кожи в период линьки и наличием подроста новых волос при сохранении еще не успевших выпасть старых. В результате плотность волосяного покрова увеличивается с 558.4 + 3.4 волоса на 4 мм2 до 708.6 + 2.0, т. е. почти на 30 %.
Интересно, что наиболее высоким оказался показатель теплопроводности у мышовок, содержащихся в неволе и погибших в октябре. Дело в том, что к этому времени у них образовывается сохраняющийся на мездре мощный подкожный слой жира, а это мешает точному определению показателя теплопроводности. Волосы таких шкурок пропитываются жиром (особенно при неизбежном нагреве на приборе), вследствие чего нарушается их укладка – они слипаются «сосульками», становятся видны участки кожи, не прикрытые волосами. Нарушение укладки волос сказывается на их пышности и приводит к исчезновению «инертного» слоя воздуха, а это искажает получаемые показатели теплопроводности испытываемых шкурок. Коэффициент теплопроводности таких шкурок оказался в среднем в 2.9 раза выше линных и в 2.3 раза выше летних шкурок.
Линька
На таежном Севере, где лесная мышовка обитает на северной периферии ареала, линька ее продолжается в течение всего лета. У взрослых она, очевидно, проходит в июне – июле (первые линяющие зверьки добыты 8.06.1968, 18.06.1982 и 19.06.1959), у молодых – в августе – сентябре. В июне линяло 12.5 % взрослых особей, в июле – 15.5, в августе – сентябре – 38.1 % прибылых.
Линное пятно появляется обычно на крестце, затем расширяется и одновременно продвигается вперед – к голове и на грудь. К моменту, когда шкурка со стороны мездры оказывается сплошь черной, на задней части спины начинают пробиваться кончики новых волос. При этом «фоновая» шерсть не выглядит тусклой и изношенной, как это обыкновенно наблюдается у землероек, кончики волос не стерты.
Указать более точные сроки линьки, порядок смены волос или половые отличия в этом процессе мы не можем, поскольку лишь немногие из 49 зверьков, отнесенных нами к группе линяющих, имели следы интенсивной линьки – обширную (на 60–100 % площади шкурки) пигментацию на мездре. Чаще же у них обнаруживались мелкие пятнышки, свидетельствующие об окончании или начале линьки. Да и общая доля таких «линяющих» зверьков среди всех добытых оказалась сравнительно невелика – у взрослых она составила 10 %, у молодых – 39 %. По всей вероятности, это связано с понижением двигательной активности животных во время линьки, с тем, что они ведут в это время скрытый образ жизни и потому редко попадаются в канавки и давилки.
В нашей коллекции нет ни одного зверька с зимним волосяным покровом, отличающимся от летнего длиной, густотой и толщиной волос. Это могло произойти по двум причинам: либо зверьки залегают в зимнюю спячку прежде, чем успевает сформироваться зимний мех, и такие особи, естественно, в ловушки и канавки не попадаются, либо появляющаяся в результате линьки зимняя шерсть не отличается от старой по морфологическим показателям. Последнее тем более вероятно, если учесть, что резкое падение метаболизма в зимний период не требует у впадающей в спячку мышовки соответствующих изменений структуры волос. Находясь в спячке, животное долгое время остается неподвижным, а это само собой исключает конфекционную теплоотдачу.
Заключение
Настоящей публикацией завершается серия наших работ, посвященных особенностям строения, теплозащитных свойств и линьки волосяного покрова различных видов мелких млекопитающих таежного Севера – кроту, землеройкам-бурозубкам, водяной куторе, рыжим и серым полевкам. Как следует из проведенных исследований, волосяной покров этих достаточно далеких друг от друга и в плане систематики (принадлежат к разным отрядам), и в экологическом отношении животных (среди них норные, наземные, полуводные виды) характеризуется не только общими, свойственными всем представителям этой группы зверьков признаками (дифференциация волос на 3 или 4 категории, трехслойность волосяного стержня, относительно небольшая длина и толщина волос, зональность окраски, соотношение слоев волоса, своеобразие сезонной изменчивости и других морфологических показателей шерсти), но и специфическими видовыми особенностями, имеющими важное приспособительное значение и представляющими поэтому особый интерес для сравнительно-экологического анализа. Несмотря на внешнее, во многом кажущееся, сходство в строении шерстного покрова, у различных исследованных видов мелких млекопитающих обнаруживаются существенные отличия и в структуре покровов, и в гистологии отдельного волоса, и в характере распределения в нем пигмента, и в густоте, длине и толщине волос, и распределении их по телу.
Например, у всех насекомоядных волосы имеют сегментированное строение, обеспечивающее укладку волос в любую сторону и способность свободно передвигаться в узких ходах нор как вперед, так и пятясь назад, тогда как у грызунов эта сегментация отсутствует, и волосы образуют «потоки» в направлении от головы к хвосту. Кроме того, если у насекомоядных волосы растут перпендикулярно к поверхности кожи и расположены поодиночке, то у грызунов они лежат наклонно и образуют группы (пучки). Имеются отличия и в микроструктуре стержня волос. Так, на граннах остевых волос землероек (в отличие от грызунов) обнаруживаются увеличивающие их прочность продольные желобки, отчего в поперечнике волос имеет вид двутаврового среза. Сердцевина волоса у насекомоядных прерывистая, однорядная, а у грызунов сплошная и в гранне 3–4-рядная.
Окраска меха определяется наличием и различной концентрацией меланина и липохрома (черного и рыжего пигментов). При этом липохром располагается в граннах, а меланин – в остальных участках волос. Общей для всех исследованных видов является темная с преобладанием бурых тонов окраска дорсальной стороны тела и светлая, грязно-белая (за исключением крота) вентральной. У большинства представителей имеется хорошо выраженная переходная зона на боках, у некоторых (кутора, крот) она отсутствует. В группе мелких грызунов особняком стоит лесная мышовка – своеобразный лесной тушканчик, единственный из всех обитающих на Севере мелких звкерьков, впадающий в зимнюю спячку.
Наконец, стала окончательно ясна причина исключительной холодостойкости мелких млекопитающих, обладающих легким, относительно тонкослойным и негустым шерстным покровом и тем не менее отличающихся исключительной устойчивостью к низким температурам. Выяснилось, что все дело в их совершенной физической терморегуляции, при которой стойкостью к холоду они обязаны насыщающему ворс инертному воздуху, лучшему в мире изолятору, успешно препятствующему потерям тепла благодаря соответствующим морфологическим особенностям волосяного покрова.
Проведенные исследования, разумеется, не претендуют на исчерпывающую полноту, но, как надеется автор, смогут всемерно помочь тем, кто решится на их дальнейшее продолжение.
Библиография
Ивантер Э. В. К экологии лесной мышовки (Sicista betulina Pall.) // Aquilo. Ser. Zool. 1972. Vol. 13. P. 103–108.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР . Л.: Наука, 1975. 247 с.
Ивантер Э. В. Основы практической биометрии . Петрозаводск: Карелия, 1979. 103 с.
Ивантер Э. В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала . М.: Товарищество научных изданий КМК, 2018. 770 с.
Ивантер Э. В. Строение, теплозащитные свойства и линька волосяного покрова обыкновенной бурозубки (Sorex araneus) // Зоологический журнал. 2021. Т. 100, № 4. С. 865–880.
Ивантер Э. В., Ивантер Т. В., Туманов И. Л. Адаптивные особенности мелких млекопитающих. Эколого-морфологические и физиологические аспекты . Л.: Наука, 1985. 318 с.
Ивантер Э. В., Коросов А. В. Основы биометрии. Введение в статистический анализ биологических явлений и процессов . Петрозаводск: Изд-во ПетрГУ, 1992. 164 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск: Изд-во ПетрГУ, 2003. 298 с.
Ивантер Э. В., Левина Р. В. Морфологическая характеристика теплозащитных свойств шерстного покрова насекомоядных млекопитающих // Вид и его продуктивность в ареале: Метериалы III Всесоюзного совещания. Вильнюс, 1980. С. 121–125.
Ивантер Э. В., Левина Р. В. Структура и теплоизоляционные свойства волосяного покрова средней бурозубки (Sorex caecutiens Laxm.) // Экология наземных позвоночных Северо-Запада СССР. Петрозаводск, 1981. С. 110–119.
Ивантер Э. В., Левина З. В. Морфология, теплозащитные свойства и линька волосяного покрова малой бурозубки // Вид и его продуктивность в ареале. М.: Наука, 1983. С. 15–26.
Ивантер Э. В., Левина Р. В., Ивантер Т. В. Адаптивные особенности структуры волосяного покрова и линьки у полуводных млекопитающих (на примере обыкновенной куторы) // Зоологический журнал. 1984. Т. 63, вып. 2. С. 245–255.
Ивантер Э. В., Моисеева Е. А. Материалы к изучению экологии средней бурозубки (Sorex caecutiens Laxm.) в европейской части ареала. Сообщение 1. Численность и биотопическое размещение // Принципы экологии. 2022. № 1. С. 47–57.
Коросов А. В. Специальные методы биометрии . Петрозаводск: Изд-во ПетрГУ, 2007. 264 с.
Кривошеев В. Г., Варич А. А., Уманцева И. Д., Кривошеева В. П. Механизмы физической терморегуляции сибирского лемминга и субарктических популяций красной полевки и полевки-экономки // Материалы по экологии мелких млекопитающих Субарктики. Новосибирск, 1975. С. 119–144.
Крыльцов А. И. Материалы по линьке мышевидных грызунов. Сообщение 1. Линька массовых видов полевок северного Казахстана // Зоологический журнал. 1958. Т. 37, вып. 2. С. 271–286.
Крыльцов А. И. Топография линьки грызунов и возможность использования ее в качестве таксономического признака // Труды Института защиты растений Казахской ССР. 1962. Т. 7. С. 418–451.
Кузнецов Б. А. Товароведение пушно-мехового сырья СССР . М.: Госиздат, 1932. 464 с.
Кузнецов Б. А. Основы товароведения пушно-мехового сырья . М.: Заготиздат, 1952. 508 с.
Марвин М. Я. Строение волосяного покрова полевок рода Microtus Среднего Урала // Ученые записки Уральского государственного университета. Серия биол. 1966. Вып. 3. С. 52–57.
Марвин М. Я. Строение волосяного покрова грызунов Урала // Научные труды Свердловского государственного педагогического института. Сборник 235. 1974. С. 11–22.
Назарова И. В. Эколого-морфологический очерк серых полевок Волжско-Камского края // Известия Казанского филиала АН СССР. Серия биол. 1958. № 6. С. 27–41.
Огнев С. И. Звери СССР и прилежащих стран . М.: Изд-во АН СССР, 1950. 706 с.
Соколов В. Е. Кожный покров млекопитающих . М.: Наука, 1973. 487 с.
Соколов В. Е., Скурат Л. Н., Степанова Л. В. и др. Руководство по изучению кожного покрова млекопитающих . М.: Наука, 1988. 279 с.
Хмелевская Н. В. Строение кутикулы волос грызунов, ее изменчивость и значение для систематики // Зоологический журнал. 1965. Т. 44, вып. 7. С. 1064–1074.
Церевитинов Б. Ф. Дифференцировка волосяного покрова пушных зверей // Труды Всесоюзного научно-исследовательского института охотничьего хозяйства. М., 1951. Вып. 10. С. 6–17.
Церевитинов Б. Ф. Топографические особенности волосяного покрова пушных зверей // Труды Всесоюзного научно-исследовательского института животного сырья и пушнины. М., 1958. Вып. 17. С. 256–307.
Bauer K. Kenntnis von Microtus oeconomus mehely Ehik // Zool. Jahrb., Abt. 1953. 3 / Bd. 82. 1–2. S. 114–126.
Благодарности
Работа проводилась в плане выполнения Государственного задания Федерального исследовательского центра «Карельский научный центр Российской академии наук».

 © 2011 - 2026
© 2011 - 2026