



Введение
В представленном сообщении подводятся итоги многолетних (1958–2019 гг.) исследований размножения, возрастно-половой структуры популяции и динамики численности лесной мышовки в условиях Восточной Фенноскандии. При этом основное внимание уделялось региональным особенностям соответствующих экологических показателей вида, и прежде всего связанным с обитанием его у северных границ распространения.
Материалы
Материалы и методы представлены в первой части (Ивантер, 2021).
Результаты
Размножение
Точное определение возраста зверьков и анализ состояния их половой системы (табл. 1, рис. 1) показывают, что в условиях таежного Северо-Запада России лесные мышовки впервые начинают размножаться в возрасте около года, после первой зимовки. Никаких изменений в гонадах сеголетков, указывающих на возможность их созревания и участия в размножении в год своего рождения, не обнаружено.
Таблица 1. Характеристика половой системы лесной мышовки
| Показатель | Зимовавшие | Прибылые, август | ||
| июнь | июль | август | ||
| Самцы | ||||
| Число исследованных зверьков | 166 | 75 | 18 | 68 |
| Длина семенника, мм | 6.7 (3.1–12.2) | 5.8 (2.5–12.1) | 4.9 (3.2–6.0) | 3.5 (1.0–5.0) |
| Длина семенных пузырьков, мм | 8.4 (3.5–11.7) | 7.2 (4.5–11.0) | 3.0 (2.5–4.5) | 2.3 (1.0–3.6) |
| Вес двух семенников, мг | 54.1 (5.6–250.0) | 150 (69.0–205.0) | 68 (33.0–130.0) | 38.7 (28.0–57.0) |
| Самки | ||||
| Число исследованных зверьков | 22 | 40 | 11 | 59 |
| Длина рогов, мм | 34.4 (11.5–53.8) | 32.3 (10.4–47.8) | 17.8 (16.1–21.6) | 19.1 (6.0–31.0) |
| Длина влагалища, мм | 3.6 (2.7–11.2) | 6.4 (1.5–17.4) | 10.1 (2.5–18.7) | 3.3 (1.0–10.5) |
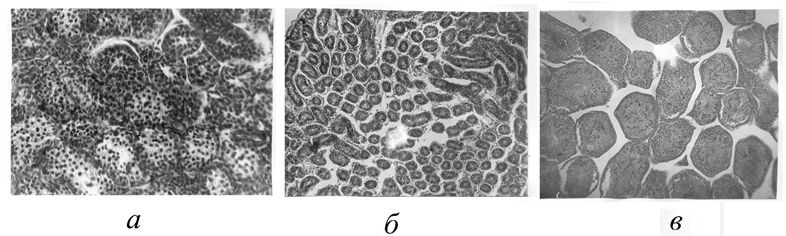
Рис. 1. Поперечные срезы извитых канальцев семенников молодого самца лесной мышовки на стадии покоя (а) и взрослого в период пассивного (б) и активного (в) сперматогенеза. Микрофото, х 400
Fig. 1. Cross sections of the convoluted tubules of the testes of a young male birch mouse at the resting stage (a) and those of an adult during the period of passive (б) and active (в) spermatogenesis. Microphoto, x 400
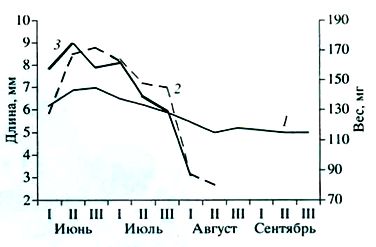
Рис. 2. Средняя длина семенника (1), семенных пузырьков (2) и средний вес двух семенников (3) у лесных мышовок по декадам
Fig. 2. Average length of testis (1), seminal vesicles (2) and average weight of two testes (3) in birch mice by decades
Размеры семенников не превышают у них 5.0 х 3.3 мм, а вес 57 мг (табл. 1, рис. 2). Инфантильная матка имеет рога до 17, влагалище до 10.5 мм. У взрослых мышовок размеры половых органов в несколько раз больше. Вес двух семенников 33–250 мг, длина семенника 3.2–12.1 мм, длина рогов матки 9.0–32.0, влагалища 2.5–18.7 мм.
Семенные пузырьки выражены только у зимовавших особей. О позднем половом созревании лесных мышовок (отсутствии размножения молодых, созревающих лишь на следующий год после рождения) сообщают и другие исследователи (Попов, 1960; Реймерс, Воронов, 1953; Штильмарк, 1965; Кулик и др., 1968; Айрапетьянц, 1969; Лямкин и др., 1983; Буйдалина, 1987; Шенброт и др., 1995).
О сроках и интенсивности размножения лесной мышовки в Карелии можно судить по результатам специального гистологического исследования половых циклов самцов и самок (рис. 1, 3). В начале июня, когда у этого вида начинается сезон репродукции, функциональное состояние половых органов самцов характеризуется активным сперматогенезом. Внешне это выражается в значительном увеличении веса семенников (к июлю до 177 мг) и их размеров (до 7.2–7.9 мм к концу июня), а также в развитии придаточных желез. В семенных канальцах (средний диаметр 192 мкм) присутствуют все клеточные элементы от клеток Сертоли и сперматогониев до зрелых сперматозоидов, собирающихся в просветах канальцев.
Гистологическая картина сперматогенеза и сезонные изменения размеров семенников (см. рис. 1, 2) показывают его стабильную активность в период с начала – середины июня и до конца июля, т. е. в течение большей части периода бодрствования, отсутствие каких-либо подъемов и спадов, соответствующих периодам спаривания и паузам между ними. Постепенное затухание сперматогенеза и дегенерация зародышевого эпителия до однорядного слоя сперматогониев и клеток Сертоли начинаются в августе, но в массе происходят, очевидно, только в сентябре, перед впадением в спячку. У молодых самцов гонады находятся в состоянии полного покоя, генеративный эпителий содержит лишь клетки Сертоли и сперматогонии (см. рис. 2).
Изучение астрального цикла самок (см. рис. 3) показало, что лесная мышовка – моноциклический вид, лишь однажды за лето приходящий в состояние эструса, причем по времени эструсы самок соответствуют активному сперматогенезу самцов. В июне половые органы самок увеличены, а в июле достигают максимальных размеров. Особенно резко возрастает вес яичников (до 10–12 мг). В яичниках много фолликулов на разных стадиях развития, от примордиальных до граафовых пузырьков, диаметром от 130 до 300 мкм. Ткань яичника богата кровеносными сосудами. Некоторые полостные фолликулы близки к разрыву. Эпителий слизистой влагалища многорядный, в верхних слоях хорошо заметно ороговение клеток, происходит их усиленная десквамация. У беременных самок (в июне – июле) яичник содержит желтые тела беременности (их максимальный диаметр 258 мкм) и атрезии. Матка гиперемирована и на поперечном разрезе имеет звездчатую форму, в просвете – секрет маточных желез. После родов рост фолликулов замедляется, прекращается он на стадии образования многослойного зачаткового эпителия; одновременно увеличивается число атрезирующих фолликулов.
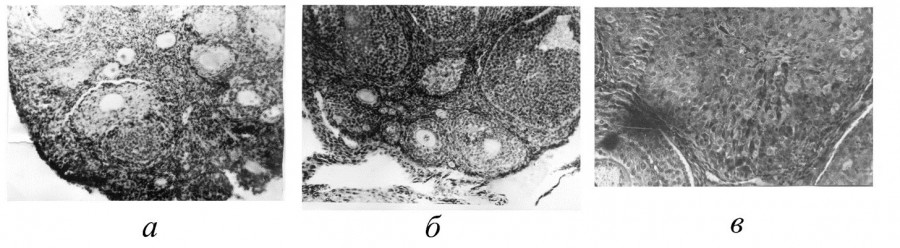
Рис. 3. Срезы половых органов взрослой самки лесной мышовки: яичник в период начала (а) и разгара (б) фолликулярной деятельности; влагалище во время эструса (в)
Fig. 3. Sections of the genital organs of an adult female wood mouse: the ovary in the period of the beginning (а) and the height (б) of follicular activity; vagina during estrus (в)
Общий ход и интенсивность размножения самок лесной мышовки по декадам представлены в табл. 2. Согласно этим данным, во второй декаде июня (до этого времени взрослые самки нам не попадались) около трети взрослых самок были беременны, остальные или еще не размножались (64 %), или уже завершили кормление недавно появившегося выводка (7 %). К концу июня число беременных (42 %), лактирующих (5 %) и рожавших (26 %) самок значительно возрастает, а без следов размножения, напротив, сокращается (26 %). В первой декаде июля более 90 % особей принимает участие в размножении, при этом процент беременных достигает максимума (60 %), затем их доля постепенно снижается. К концу месяца большинство самок уже обзаводится потомством (78 %) и либо приступает к лактации (43 %), либо уже откормило выводок (36 %). В августе отлавливались только кормящие самки (32 %), остальные уже закончили размножение. Все это говорит о том, что в условиях Карелии лесные мышовки приносят за лето только один выводок (об этом говорит и одновершинный характер кривой уловов молодых), причем массовое рождение детенышей приходится на первую половину июля.
Таблица 2. Интенсивность размножения взрослых самок лесной мышовки (сводные данные за 1966–2017 гг.)
| Месяц и декада | Число | Количество самок (% от общего числа) | |||
| исследованных | |||||
| самок | беременных | кормящих | рожавших | без следов размножения | |
| Июнь (всего) | 33 | 36.4 | 3 | 18.2 | 42.4 |
| II | 14 | 28.6 | – | 7.1 | 64.3 |
| III | 19 | 42.1 | 5.3 | 26.3 | 26.3 |
| Июль (всего) | 67 | 40.3 | 17.9 | 23.9 | 17.9 |
| I | 25 | 60 | 4 | 28 | 8 |
| II | 28 | 39.3 | 17.9 | 14.3 | 58.6 |
| III | 14 | 7.1 | 42.9 | 35.7 | 14.3 |
| Август (всего) | 22 | – | 31.8 | 31.8 | 36.4 |
| I | 9 | – | 55.6 | 33.3 | 11.1 |
| II | 8 | – | 25 | 50 | 25 |
| III | 5 | – | – | – | 100 |
| Сентярь (всего) | 8 | – | – | – | 100 |
Если учесть, что беременность длится у этого вида 25–30 дней (Godfrey, 1955, цит. по: Kubik, 1952), картина размножения лесной мышовки в Карелии представляется следующим образом. Массовый гон и спаривание происходят в первой половине июня, рождение детенышей – в первой – второй декадах июля, а их массовое расселение (переход к самостоятельной жизни и выход из гнезд) – в августе, т. е. через 30–40 дней после рождения. Это соответствует продолжительности гнездового развития (35–37 дней), установленной в результате прямых наблюдений за жизнью новорожденных (Айрапетьянц, 1969; Фокин, 1978).
Обсуждаемые ниже сезонные изменения численности мышовок свидетельствуют о сравнительно позднем их размножении в Карелии. Вследствие этого здесь наблюдаются и более поздние выход и расселение молодых особей (не ранее последней декады июля). В более южных районах это происходит почти на месяц раньше (Kubik, 1952; Попов, 1960; Кулик и др., 1968; Цветкова, 1978). Лишь в средней Сибири размножение этого вида проходит в близкие с Карелией сроки (Реймерс, 1966; Буйдалина, 1987; Лямкин и др., 1983).
Сроки размножения лесной мышовки меняются по годам в зависимости от метеорологической обстановки и, прежде всего, от хода весны. При раннем наступлении весенних явлений, теплой и сухой погоде в мае – июне размножение зверьков начинается раньше и проходит в более сжатые сроки. Напротив, в годы с поздней, затяжной и холодной весной размножение сильно задерживается и растягивается. Наиболее раннее начало размножения лесной мышовки наблюдалось в Карелии в 1959 г., когда переход среднесуточной температуры через 0 и 5 °С был отмечен 13 и 26 апреля соответственно. Гон у мышовок проходил в тот год в начале июня, первые беременные самки встречены 7 и 9 июня, а первые молодые – в конце июля (26.07). Почти на полмесяца позднее началось размножение в 1951 и 1958 гг. Переход среднесуточной температуры через 0 и 5 °С в 1951 г. произошел 25 апреля и 19 мая, в 1958 г. – 28 апреля и 22 мая. В связи с этим задержалось и размножение. Первые беременные самки были добыты в самом конце июня (27.06.1951 г. и 25.06.1958 г.), первые молодые – во второй декаде августа (12.08.1951 г. и 18.08.1958 г.).
Гораздо стабильнее интенсивность размножения (табл. 3). Доля участвующих в нем перезимовавших самок почти не меняется по годам и составляет 98–100 %. Несущественны различия и в разовой плодовитости. Таким образом, в отличие от ряда других видов мелких млекопитающих Карелии, годовые изменения численности лесной мышовки обусловлены в основном экзогенными факторами. Размножение этого вида характеризуется невысоким темпом (размножаются только взрослые особи и всего 1 раз в год) и поэтому не может служить эффективным регулятором его численности.
С другой стороны, популяция мышовок в Карелии отличается общей невысокой плотностью и даже в годы пиков численности не достигает критической величины. В этих условиях вряд ли есть необходимость в выработке внутрипопуляционных механизмов автоконтроля. Их отсутствие является следствием общей невысокой численности вида у северной границы ареала и одновременно причиной резких колебаний его плотности на этом фоне. Не исключено, что в отдельных участках ареала с оптимальными условиями, где средний многолетний уровень численности вида в несколько раз выше, чем на Северо-Западе России, механизмы эндогенной саморегуляции все же имеют место. Иначе трудно объяснить стабильность численности популяции при высоком уровне ее плотности и отсутствие этой стабильности при низкой плотности популяции. В первом случае численность может регулироваться за счет изменений числа участвующих в размножении взрослых самок (в Татарстане и Кировской области в некоторые годы отмечалось до 10 и даже до 70 % яловых самок (Попов, 1960; Кулик и др., 1968)), а во втором с этой задачей вполне справляются внешние факторы (например, климатические условия), и интенсивность размножения не меняется по годам.
Средний размер выводка у лесных мышовок Карелии на основании подсчета эмбрионов у 53 беременных самок составил 5.1 ± 0.2 с колебаниями от 3 до 11 (табл. 4). Это несколько ниже, чем в оптимуме ареала – в Кировской области, на юге Коми и Татарстане, где средний показатель плодовитости равнялся соответственно 5.5, 5.9 и 5.5 (в Волжско-Камском заповеднике, по данным Назаровой и Мовчаренко (1980), эта величина составила 6.4), но выше, чем в предгорьях Алтая (4.3 эмбриона на самку), долине р. Верхняя Ангара (3.7) и Ивановской области (4.0) (Попов, 1960; Кулик и др., 1968; Лямкин и др., 1983; Буйдалина, 1987; Хелевина, Окулова, 1988).
Среднее число эмбрионов, приходящееся на одну самку, в течение репродуктивного периода практически не меняется. Наибольшей плодовитостью отличаются первородящие годовалые самки; с возрастом она снижается. Различий между средним числом эмбрионов и плацентарных пятен, на что указывал для Татарстана В. А. Попов (1960), на нашем материале не выявлено: на одну беременную самку приходится в среднем 5.11 + 0.30 эмбриона, а на одну рожавшую – 5.16 + 0.32 плацентарных пятна.
Обращает внимание весьма значительная асимметричность кривой распределения самок по числу эмбионов в выводке (показатель «косости» А равен +1.5, т = 0.39, t = 4.4). Это указывает на существование в популяции лесных мышовок Карелии отчетливой тенденции к увеличению разовой плодовитости и свидетельствует о том, что процесс эволюционного отбора на повышенную плодовитость (призванную компенсировать повышенную в периферических популяциях смертность) не завершен: уровень плодовитости еще не достиг оптимального для данных условий. Мышовка проникла на север сравнительно недавно, и адаптация ее к местным условиям, по-видимому, продолжается.
Таблица 3. Интенсивность размножения взрослых самок лесной мышовки (сводные данные за 1966–2017 гг.)
| Месяц и декада | Число | Количество самок (% от общего числа) | |||
| исследованных | |||||
| самок | беременных | кормящих | рожавших | без следов размножения | |
| Июнь (всего) | 33 | 36.4 | 3 | 18.2 | 42.4 |
| II | 14 | 28.6 | – | 7.1 | 64.3 |
| III | 19 | 42.1 | 5.3 | 26.3 | 26.3 |
| Июль (всего) | 67 | 40.3 | 17.9 | 23.9 | 17.9 |
| I | 25 | 60 | 4 | 28 | 8 |
| II | 28 | 39.3 | 17.9 | 14.3 | 58.6 |
| III | 14 | 7.1 | 42.9 | 35.7 | 14.3 |
| Август (всего) | 22 | – | 31.8 | 31.8 | 36.4 |
| I | 9 | – | 55.6 | 33.3 | 11.1 |
| II | 8 | – | 25 | 50 | 25 |
| III | 5 | – | – | – | 100 |
| Сентярь (всего) | 8 | – | – | – | 100 |
Таблица 4. Вариации величины выводка у лесной мышовки в Карелии
| Месяц | Общее число беременных самок | Количество самок (экз.) с числом эмбрионов | Среднее число эмбрионов на самку | ||||||||
| 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||
| Июнь | 12 | 2 | 3 | 5 | – | 1 | – | – | – | 1 | 5.1 (2.2 + 2.9) |
| Июль | 35 | 2 | 13 | 7 | 9 | 3 | – | 1 | – | – | 5.1 (2.6 + 2.5) |
| Август | 6 | – | 1 | 3 | 2 | – | – | – | – | – | 5.2 (2.8 + 2.4) |
| Всего | 53 | 8 | 21 | 20 | 11 | 4 | – | 1 | – | 1 | 5.1 (2.6 + 2.5) |
| Из них: самки в возрасте 1 года | |||||||||||
| Июль | 21 | 1 | 4 | 8 | 6 | 1 | – | 1 | – | – | 5.3 (2.9 + 2.4) |
| Самки в возрасте 2 лет и старше | |||||||||||
| Июль | 8 | 2 | 5 | – | 1 | – | – | – | – | – | 4.0 (2.0 + 2.0) |
Примечание. В скобках указано число эмбрионов в правом и левом рогах матки.
Экологическая структура популяции
Половой состав отловленных взрослых мышовок в значительной степени отражает изменение активности зверьков разного пола в связи с размножением (табл. 5, рис. 4, 5). В первой декаде июня ловятся исключительно самцы, во второй половине месяца они составляют около 90 %, а затем, обычно ко второй половине июля, по мере снижения половой активности самцов и освобождения самок от забот о выводке, соотношение полов выравнивается. В августе оно снова уклоняется от 1:1, но теперь в пользу самок (70.6 %). К этому времени они уже откормили выводок, и активность их, а вместе с ней и отлов увеличиваются.
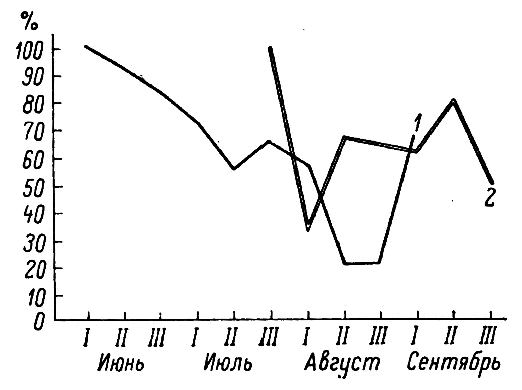
Рис. 4. Изменение доли (%) самцов у взрослых (1) и молодых (2) лесных мышовок по декадам
Fig. 4. Change in the proportion (%) of males in adults (1) and young (2) birch mice by decades
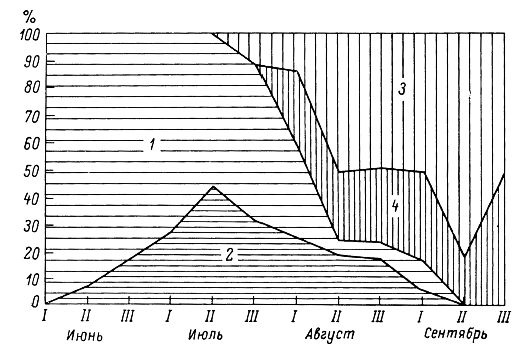
Рис. 5. Изменение возрастно-половой структуры (доля в уловах, %) лесной мышовки (сводные данные отлова за все годы исследований): 1 – зимовавшие самцы; 2 – зимовавшие самки; 3 – прибылые самцы; 4 – прибылые самки
Fig. 5. Changes in the age-sex structure (share in catches, %) of the birch mouse (summary data of trapping for all years of research): 1 – overwintered males; 2 – overwintered females; 3 – arrived males; 4 – arrived females
Таблица 5. Возрастной и половой состав популяции лесной мышовки в Карелии (по данным отлова
в 1959–2016 гг.)
| Месяц | Число зимовавших | Число прибылых | Общее число зверьков | |||||
| всего | самцы, % | ᵪ2 | всего | самцы, % | ᵪ2 | всего | зимовавшие, % | |
| Июнь | 275 | 88.4 | 162 | – | – | – | 275 | 100 |
| Июль | 171 | 58.5 | 4.9 | 12 | 58.3 | 0.3 | 183 | 93.4 |
| Август | 33 | 36.4 | 2.5 | 145 | 53.1 | 0.6 | 178 | 18.5 |
| Сентябрь | 3 | 66.6 | 0.3 | 30 | 63.3 | 2.1 | 33 | 9.1 |
| Всего | 482 | 74.1 | 111.7 | 187 | 55.1 | 1.9 | 669 | 72 |
Общая доля самцов в уловах взрослых зверьков составила по всему материалу 74.1 %, причем сопоставление полученных величин хи-квадрат с критической (3.84) говорит о большой статистической достоверности отмеченной диспропорции. Приблизительно такая же картина наблюдается и в других частях ареала (Снигиревская, 1954; Попов, I960; Плешак, Козловский, 1983; Буйдалина, 1987; Kubik, 1952 и др.).
В уловах молодых мышовок с июля по сентябрь также преобладали самцы, но их количественное доминирование было не столь существенно. Тем не менее высказанное нами ранее (Ивантер, 1972, 1975) предположение о том, что это связано не с большей их активностью, а с количественным преобладанием в популяции вследствие более частого рождения особей мужского пола, остается в силе. К близкому выводу приходят и другие исследователи (Попов, I960; Реймере, Воронов, 1963; Лямкин и др., 1983). Это позволяет отнести лесную мышовку к видам со стойким численным преобладанием самцов (Кубанцев, 1972; Большаков, Кубанцев, 1984).
Возрастной состав популяции мышовок меняется по сезонам (табл. 6, рис. 5). До конца июля в ловушки и канавки попадают почти исключительно взрослые перезимовавшие особи, а начиная с первой декады августа все больший удельный вес в уловах занимают молодые зверьки. В начале месяца их не более 40 %, в середине и конце месяца 84 %, а в сентябре 90–100 %. Ко второй декаде сентября взрослые мышовки почти полностью исчезают, и ловятся только прибылые зверьки, родившиеся в текущем году. При этом сокращается не только относительная (доля в уловах), но и абсолютная численность взрослых животных. Такое резкое изменение возрастной структуры популяции мышовок в конце лета связано как с процессами естественной смертности взрослых особей, так и с более ранним уходом их на зимовку. Последнее характерно, по-видимому, и для зверьков других популяций (Кулик и др., 1968; Фокин, 1978).
Сроки появления молодых мышовок в отловах варьируют в разных частях ареала. В долине р. Верхняя Ангара они попадались с конца июля (Лямкин и др., 1983), в тайге Сосьвинского Приобья – несколько позже (в начале августа), причем особенно резкое увеличение их количества в отловах произошло в третьей декаде этого месяца. В южных же частях ареала (Татарстан, Оренбургская область, предгорья Алтая) прибылые зверьки начинают встречаться почти на месяц раньше, с первых чисел июля (Попов, 1960; Кулик и др., 1968; Цветкова, 1978).
Большой интерес представляет анализ возрастного состава взрослой части популяции (см. табл. 6). Благодаря применению специальной методики определения возраста мышовок (по слоистым структурам кости нижней челюсти) удалось установить, что основу группы зимовавших зверьков составляют особи, родившиеся в прошлом году (80 %), далее следуют мышовки, пережившие две зимы (16 %), и совсем редки особи трех лет и старше (4 %). В августе – сентябре, перед уходом на зимовку, популяция имеет следующую структуру: прибылые составляют около 80 %, годовики – 16.4, двухлетки – 2.7, трехлетние и более старые – 0.9 %.
Таким образом, численность каждой следующей возрастной группы сокращается по сравнению с предыдущей в 5–6 раз, или в среднем на 80 %. Эти 80 % и составляют величину годовой смертности населения мышовок, причем, судя по тому, что количественное соотношение между соседними возрастными группами довольно стойко сохраняется, темп смертности у них приблизительно одинаков.
Таблица 6. Возрастная структура популяции лесной мышовки (но исследованию черепов)
| Возрастные группы | По слоистым структурам | По стертости зубов | ||
| n | % | n | % | |
| Сеголетки | 49 | 32.9 | 76 | 45.5 |
| Годовики | 80 | 53.0 | 66 | 39.5 |
| Двухлетки | 16 | 10.7 | 24 | 14.4 |
| Трехлетки | 3 | 2.0 | 1 | 0.6 |
| Четыре года и старше | 1 | 0.7 | 0 | 0 |
| Всего | 149 | 100 | 167 | 100 |
Итак, весной популяция лесных мышовок на таежном Северо-Западе России состоит из перезимовавших зврьков, из которых 81.8 % – родившиеся в прошлом году, 13.6 % – пережившие две зимы и 4.5 % – особи 3 лет и старше. За лето доля перезимовавших зверьков сокращается, и популяция на 80 % состоит из молодых зверьков. Это близко к данным по бассейну средней Вычегды (Куприянова, 1994), где в августе население мышовок на 17.5 % состот из перезимовавших особей и на 82.5 % – из прибылых. К концу августа мышовки значительно реже начинают ловиться здесь в канавки и в сентябре залегают в спячку.
Соотношение же полов как среди зимовавших, так и у сеголеток в августе сильно сдвинуто в пользу самцов. Так, в бассейне средней Вычегды среди отловленных зверьков (n = 274) самки сеголетки составляли 34.7 %, самцы – 47.8 %, перезимовавшие – 5.1 и 12.4 % соответственно. Стойкое преобладание самцов характерно для лесной мышовки и в других частях ареала (Снигиревская, 1954; Попов, 1960; Kubik, 1952). По нашему мнению, о чем мы неоднократно высказывались ранее (Ивантер, 1975; Ивантер, Кухарева, 2008), это, скорее всего, связано с более частым рождением особей мужского пола.
Динамика численности
В среднем для всех охваченных учетами районов Южной Карелии показатели численности лесной мышовки в расчете на 10 канавко-суток изменялись за годы исследований (1959–2014 гг.) в 15–25 раз (рис. 6): от 0–0.1 (1964, 1966, 1974–1976, 1979, 1982, 2004 гг.) до 1.7–2.4 (1970, 1974, 1980, 1986, 1996, 1998, 2000–2002 гг.). За смежные годы наблюдались более умеренные 4–9-кратные изменения. Таким образом, в южной части Карелии, расположенной у северной границы области распространения вида, численность лесной мышовки отличалась крайней нестабильностью и гораздо большей изменчивостью, чем в большинстве других мест ареала, например в Предгорьях Урала, Средней Сибири, Кировской и Ивановской областях (Даргольц, 1964; Кулик и др., 1968; Хелевина, Окулова, 1988), где амплитуда колебаний не превышала 3–12 раз. В то же время в Татарстане, который, как и Карелия, представляет окраину ареала вида (в данном случае южную), В. А. Попов (1960) на протяжении 11 лет наблюдал 10–12-кратные изменения его численности (от 0.2 до 2.1 экз. на 10 канавко-суток). Это соответствует известному представлению о том, что численность вида наиболее высока и устойчива в зону оптимума, а при продвижении к периферии ареала становится более изменчивой: амплитуда колебаний ее резко возрастает (Фалькенштейн, 1939; Наумов, 1945; Новиков, 1956; Ходашева, 1966 и др.).
Четкой цикличности (в смысле правильных осцилляций) в колебаниях численности лесной мышовки Карелии не обнаружено. За 50 лет стационарных наблюдений подъемы численности (до 0.8–2.5 особи на 10 канавко-суток) происходили 11 раз: в 1962, 1970, 1974, 1977, 1980, 1986, 1988, 1992, 1996, 1998 и 2000–2001 гг., т. е. через 1–7 лет (рис. 6).
Вместе с тем, если рассматривать приведенные выше данные отлова мышовки в 1951 г. как свидетельство резкого подъема ее численности, то можно, по-видимому, говорить о 10–11-летней периодичности в изменениях численности. Однако этот вывод нуждается в дальнейшем подтверждении.
Строгой периодичности в многолетних изменениях численности мышовки на северном пределе ареала не смог выявить и предпринятый нами автокорреляционный анализ (табл. 7). Рассчитанные по данным отловов ловушками и канавками коэффициенты корреляции при всех лагах не превышали минимального порогового значения, что подтверждало бы достоверность их отличия от 0. Но, с другой стороны, слабая регулярность колебаний все же имела место, о чем свидетельствуют коэффициенты корреляции, приближающиеся по величине к критическому уровню при обоих видах учетов. Так, для отловов канавками максимальное положительное значение этого показателя оказалось равным 0.28 при L = 1 (пороговое г = 0.325), что говорит о сходстве уровня численности в смежные годы, а вернее, об отсутствии резких ее перепадов в текущий и следующий за ним год. Возможно, это объясняется тем, что учеты канавками отражают не только численность, но и подвижность зверьков (Наумов, 1955), а она изменяется не так резко, как первая. При учетах давилками мы обычно фиксируем более резкие, чем при отлове канавками, колебания численности мышовок за смежные годы. Это, вероятно, связано с плохой отлавливаемостью их ловушками, отчего при подъеме численности популяции зверьки, хотя и редко, но все же попадаются в них, тогда как при снижении совсем исчезают из отловов (рис. 6). При учетах ловушками близким к достоверно отличающемуся от нуля является значение коэффициента корреляции при L, равном 5 (г = 0.31, при пороговом значении 0.35) (табл. 7), что свидетельствует о тенденции к пятилетней периодичности динамики численности, которая, впрочем, не проявляется при учетах канавками.
Гармонический анализ позволяет более детально и содержательно охарактеризовать исследуемое явление (в данном случае – многолетнюю динамику численности лесной мышовки) путем разложения исходного ряда на компоненты с разными периодами (рис. 7). При этом и канавочные, и ловушечные учеты показывают одинаково неопределенную картину: амплитуда гармоник более или менее равномерно распределена по всем частотам (периодам), что указывает на отсутствие четкой ритмичности в изменениях численности изучаемого вида. Напомним, что при строгой периодичности (которая, кстати, встречается в природе крайне редко) на периодограмме должны наблюдаться один или два высоких пика при значении амплитуд остальных гармоник, близких к 0.
При отловах канавками максимальный и значимый пик оказался равным периоду 5.4 года, что говорит об изменениях численности лесной мышовки с интервалом 5–6 лет: подъемы зафиксированы в 1974, 1980, 1985 (1986) гг., а также в 1996 и 2001 гг. (рис. 7). Этот вывод подтверждают наблюдения Назаровой и Мовчаренко (1980), согласно которым в Волжско-Камском крае изменения численности лесной мышовки происходят с периодичностью в 5–7 лет. Спектральный анализ позволил также выявить наличие в динамике численности этого вида в Карелии слабую периодичность в 12–13 лет (дисперсия гармоники с периодом 12.7 года оказалась близкой к значимой), проявившуюся в подъемах численности мышовки в 1975 (1974), 1988 и 2001 гг. (см. рис. 7). Заключение о наблюдающейся в отловах ловушками (но не регистрируемой канавочными учетами) цикличности с периодом в 2.5 года (т. е. в 2–3 года) основывается на том, что численность в 1972, 1974, 1977, 1979, 1987, 1989, 1992 и 1994 гг. попеременно принимала значения, близкие или равные 0.04 и 0 экз. на 100 ловушко-суток (см. рис. 7). Других регулярных компонент в динамике численности вида не обнаруживается.
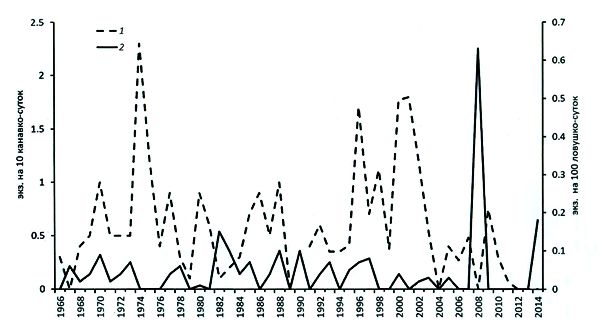
Рис. 6. Изменения численности лесной мышовки по данным учетов в Приладожье: 1 – ловушко-линии, экз. на 100 ловушко-суток; 2 – ловчие канавки, экз. на 10 канавко-суток
Fig. 6. Changes in the abundance of the birch mouse according to the counts in the Ladoga area: 1 – trap lines, specimens for 100 trap-days; 2 – trapping grooves, specimens for 10 groove days
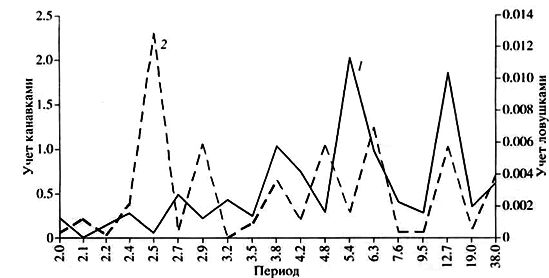
Рис. 7. Периодограмма многолетней динамики численности лесной мышовки: 1 – по данным отловов канавками,
2 – ловушками
Fig. 7. Periodogram of long-term dynamics of the birch mouse population: 1 – according to the data of catches by grooves,
2 – by traps
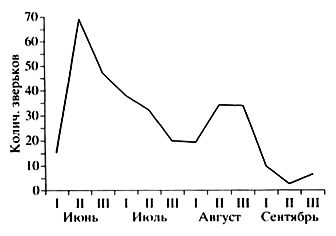
Рис. 8. Динамика отлова лесных мышовок по декадам (сводные данные); по оси абсцисс – десятилетия, по оси ординат – количество животных
Fig. 8. Dynamics of catching birch mice by decades (summary data); on the abscissa – decades, on the ordinate – the number of animals
Таблица 7. Значения коэффициентов автокорреляционной функции многолетней динамики численности лесной мышовки
| Лаг | Учеты | Пороговые значения г | |
| ловушками | канавками | ||
| 1 | -0.032 | 0.280 | 0.325 |
| 2 | -0.027 | -0.170 | 0.325 |
| 3 | 0.042 | -0.161 | 0.325 |
| 4 | -0.174 | -0.009 | 0.325 |
| 5 | 0.314 | 0.020 | 0.349 |
| 6 | -0.091 | 0.020 | 0.349 |
| 7 | -0.067 | -0.097 | 0.349 |
| 8 | -0.023 | -0.080 | 0.361 |
| 9 | -0.213 | -0.119 | 0.361 |
| 10 | 0.111 | -0.087 | 0.367 |
| 11 | -0.191 | 0.017 | 0.374 |
| 12 | 0.136 | -0.027 | 0.381 |
Сравнение хода многолетних изменений численности у разных видов мелких млекопитающих Карелии показывает, что у мышовок эти изменения происходят синхронно с таковыми у землероек, а не у полевок, численность которых колеблется, очевидно, под влиянием других факторов. Аналогия в характере многолетней динамики позволяет предположить общую причину. Как уже было показано (Формозов, 1948; Ивантер, 1975; Ивантер, Макаров, 2001), численность землероек определяют в основном условия перезимовки – глубина промерзания почвы зимой, зависящая от высоты снежного покрова и температуры воздуха. По-видимому, те же факторы действуют и на популяцию лесных мышовок. В годы, когда почва лучше защищена снегом и слабо промерзает, численность зверьков выше, чем после зим с низким снежным покровом. Например, наиболее высокая численность мышовок наблюдалась в Карелии в 1962, 1969, 1970, 1972, 1974, 1986, 1996 и 2000–2001 гг., когда средняя минимальная температура почвы в предшествующие зимы не опускалась ниже -4 °С и составляла в среднем -2 °С. Депрессия же численности отмечена в годы (1963–1966, 1979 и 2004 гг.) со средней минимальной температурой почвы от -5 до -6 °С. Таким образом, в Карелии, как и в Костромской области и Татарстане (Формозов, 1948; Попов, 1960), численность мышовок зависит от глубины промерзания почвы и смертности зверьков во время зимней спячки. Вторым важным фактором В. А. Попов (1960) считает метеорологические условия первой половины вегетационного периода. В Карелии связь между численностью мышовок и средней температурой воздуха в мае – июне также прослеживается, но она менее отчетлива, чем зависимость от условий зимовки.
О сезонных изменениях численности лесной мышовки в Карелии можно судить по динамике отлова этих зверьков в канавки и ловушки (рис. 8). Кривая отлова, отображающая вместе с численностью активность (подвижность) животных, имеет два довольно отчетливых пика. Первый приходится на вторую половину июня – начало июля, а второй – на вторую – третью декады августа. Июньский подъем связан, очевидно, с гоном (взрослые самцы составляют 90 % от пойманных в это время зверьков), а августовский – с массовым расселением прибылых мышовок (84–100 % в уловах). Такой же двухвершинный характер имеют кривые отлова в других частях ареала вида, например в предгорьях Алтая, в Кировской области и в Татарстане, но там пики смещены на более ранние сроки – третью декаду мая и вторую половину июля (Попов, 1960; Кулик и др., 1968). Это свидетельствует о более позднем начале размножения мышовок в Карелии: разница составляет в среднем около месяца. При этом в годы с ранней весной первый пик активности наблюдается на две – три недели раньше, чем в годы с холодными, поздними и затяжными веснами.
Данные о численности мышовок и ее сезонных и годовых изменениях в других частях ареала чрезвычайно ограничены, а часто и неточны, поскольку эти зверьки редко попадаются в давилки, а канавки применяются не всегда. Во многих работах отмечаются лишь единичные встречи вида при достаточно широком его распространении. Единственное исключение представляет разве что средняя тайга европейского Северо-Востока России (юг Архангельской области, леса Республики Коми и др.), где благодаря обстоятельным работам И. Ф. Куприяновой (Куприянова, 1978, 1994; Куприянова, Наседкина, 1983; Куприянова, Наумов, 1986; Куприянова и др., 1988) этот пробел успешно восполнен.
Заключение
Отмеченные выше экологические особенности, свойственные лесной мышовке в специфических условиях северо-западной периферии ареала, отличаются значительной лабильностью и гармоничной согласованностью как с плотностью и состоянием популяции, так и с внешними факторами среды. Разнообразие экологических приспособлений, их трансформация и смена во времени и пространстве, адаптивный динамизм и ориентированность на конкретную экологическую ситуацию, складывающуюся из взаимодействия экзогенных и эндогенных факторов, обуславливают относительную стабильность и оптимальное состояние популяций рассматриваемого вида в экстремальных для него приграничных условиях. Вместо узких специальных приспособлений индивидуального ранга в периферических его населениях успешно реализуются широкие структурно-популяционные адаптивные комплексы, отличающиеся динамичностью и высокой скоростью компенсаторной перестройки. Все это вплотную приближает нас к разработке общей концепции периферических популяций мегаареальных политипических видов животных. Проведенный анализ закономерных географических отличий, характеризующих популяционную специфику репродукции зверьков этого вида, обитающих на северных пределах области распространения, подтвердил выдвинутое нами ранее положение, согласно которому в экологическом центре (в оптимуме) видового ареала население вида благодаря функционированию внутрипопуляционных компенсаторных механизмов отличается не только относительно высоким уровнем численности, но и большей устойчивостью, тогда как у северных границ ареала эти стабилизирующие механизмы практически отсутствуют, и организация населения изменяется в широком диапазоне и с большей амплитудой. Соответственно более выражены и гораздо рельефнее проявляются близ северных границ распространения и специфические структурно-популяционные адаптации, направленные на максимально эффективное воспроизводство видового населения, способное обеспечить виду и его популяциям необходимое преодоление экстремальных условий экологического пессимума и в конечном счете содействовать как более широкой их экспансии за границы ареала, так и успешному закреплению в новых областях.
В общем же участие в размножении и половое созревание лесных мышовок в Восточной Фенноскандии определяются состоянием популяции в предшествующие годы и в данный период времени, совокупным действием погодных условий и биологической спецификой сезонных генераций. В разные фазы репродуктивного цикла действие каждой группы факторов неоднозначно. Условия существования лесной мышовки в изученном регионе далеки от оптимальных, о чем свидетельствует относительно низкий уровень ее численности и высокая смертность зверьков как в осенне-зимний (спячка), так и в летний период. В экологическом центре (оптимуме) ареала, где емкость угодий достаточно высока, в отдельные годы у этого вида также наблюдаются спады численности, но там они бывают редко, тогда как в Карелии, в условиях северной периферии ареала, это достаточно частое явление.
В начале сезона размножения численность перезимовавших зверьков и их осенне-зимняя выживаемость определяются в основном погодными условиями. Даже в годы, когда поголовье мышовок заметно возрастает к осени, зиму переживает только его треть. Кроме того, не только при низком, но и при достаточно высоком репродуктивном потенциале во все летние месяцы численность видового населения из-за высокой летней смертности прибылых зверьков не может достичь высокого уровня. В других популяциях этого вида регулирование численности осуществляется в большей мере через изменение доли участвующих в размножении более молодых животных.
Во второй половине сезона размножения (после сезонного пика) стратегия популяции заключается в снижении скорости полового созревания годовалых мышовок и формировании группы зверьков, уходящих в зиму и служащих основой нового периода репродуктивного цикла. Вместе с тем на северной периферии ареала особое значение в этот период приобретает ландшафтная гетерогенность территории, выполняющая роль главного стабилизирующего фактора, поддерживающего оптимальный уровень и темпы воспроизводства популяции.
Что же касается численности лесных мышовок, обитающих вблизи северных границ ареала, то она находится здесь под контролем большого числа факторов, среди которых наиболее существенны демографический состав популяции в предшествующий и настоящий моменты времени, состояние популяций совместно обитающих видов, а также погодные условия зимнего периода. Иерархия их вкладов в изменения численности и экологической структуры популяции в основном определяется качеством местообитаний, положением популяции в ареале вида и зимними, а также весенними и раннелетними условиями погоды. При этом наиболее велика зависимость численности мышовок Карелии от глубины промерзания почвы зимой и смертности зверьков во время зимней спячки.
Библиография
Айрапетьянц А. Э. К биологии лесной мышовки в Ленинградской области // Вопросы экологии и биоценологии. Л.: ЛГУ, 1969. Вып. 9. С. 115–124.
Большаков В. Н., Кубанцев Б. С. Половая структура популяций млекопитающих и ее динамика . М.: Наука, 1984. 232 с.
Буйдалина Ф. Р. К экологии лесной мышовки // Зоологический журнал. 1987. Т. 87. Вып. 4. С. 476–493.
Даргольц В. Г. Особенности динамики численности лесной мышовки в подзоне южной тайги // Современные проблемы изучения динамики численности популяций животных. М.: Наука, 1964. С. 76–84.
Ивантер Э. В. К экологии лесной мышовки (Sicista betulina Pall.) // Aquiloo. Ser. Zool. 1972. Vol. 13. P. 103–108.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР . Л.: Наука. Ленингр. отд-ние, 1975. 246 с.
Ивантер Э. В., Кухарева А. В. К экологии лесной мышовки (Sicista beyulina) на северном пределе ареала // Зоологический журнал. 2008. Т. 87. № 4. С. 476–493.
Ивантер Э. В., Макаров А. М. Территориальная экология землероек-бурозубок (Insectivora, Sorex) . Петрозаводск: ПетрГУ, 2001. 272 с.
Ивантер Э. В. К популяционной экологии лесной мышовки (Sicista betulina Pall.) на северном пределе ареала. Сообщение I. Численность, биотопическое размещение, суточная активность, питание // Принципы экологии. 2021. № 1. С. 18–29. DOI: 10.15393/j1.art.2021.11642
Кубанцев Б. С. О половом составе популяций млекопитающих // Журнал общей биологии. 1972. Т. 33. Вып. 2. С. 109–114.
Кулик Н. Л., Тупикова Н. В., Никитина Н. А., Карасева Е. В., Суворова Л. Г. Материалы по экологии лесной мышовки (Sicista betulina Pallas) // Cборник трудов Зоологического музея МГУ. 1968. Т. 10. С. 146–159.
Куприянова И. Ф. Биология и межвидовые отношения мелких лесных млекопитающих средней тайги Европейского Севера СССР : Автореф. дис. … канд. биол. наук. М., 1978. 16 с.
Куприянова И. Ф. Биотопические группировки мелких млекопитающих и динамика их численных взаимоотношений на юге Архангельской области // Фауна и экология позвоночных животных. М.: Наука, 1978. С. 114–130.
Куприянова И. Ф. Мышевидные грызуны // Фауна европейского Северо-Востока России. Млекопитающие. СПб.: Наука, 1994. Т. 2. Ч. 1. С. 68–99.
Куприянова И. Ф., Недосекина И. Б., Большаков Н. М. Влияние вырубок на население полевок рода Clethrionomys в различных регионах Средней тайги Европейского севера // Грызуны: Тез. докл. VII Всесоюзн. совещ. Л.: Наука, 1988. Т. ΙΙΙ. С. 27–28.
Куприянова И. Ф., Наумов С. П. Особенности структуры населения мелких млекопитающих Европейской тайги // Зоологический журнал. 1986. Т. 63. Вып. 11. С. 1682–1692.
Лямкин В. Ф., Малышев Ю. С., Пузанов В. М. К экологии лесной мышовки на северо-восточной оконечности ареала // Грызуны: Материалы VI Всесоюзн. совещ. Л.: Наука, 1983. С. 328–329.
Назарова И. В., Мовчаренко В. В. Лесная мышовка в Волжско-Камском заповеднике // Грызуны: Материалы V Всесоюзн. совещ. М.: Наука, 1980. С. 242–244.
Наумов Н. П. Географическая изменчивость динамики численности животных и эволюция // Журнал общей биологии. 1945. Т. 6. Вып 1. С. 112–127.
Новиков Г. А. Еловые леса как среда обитания и роль в их жизни млекопитающих и птиц // Роль животных в жизни леса. М., 1956. 289 с.
Плешак Т. В., Козловский И. С. К экологии лесной мышовки // Биология и промысел охотничьих животных. Пермь, 1983. С. 57–61.
Попов В. А. Млекопитающие Волжско-Камского края . Казань, 1960. 466 с.
Реймерс Н. Ф. Птицы и млекопитающие южной тайги Средней Сибири . М.; Л.: Наука, 1966. 419 с.
Реймерс Н. Ф., Воронов Г. А. Насекомоядные и грызуны Верхней Лены . Иркутск, 1963. 180 с.
Снигиревская Е. М. Экология и хозяйственное значение мышевидных грызунов в широколиственных лесах Жигулевской возвышенности : Автореф. дис. … канд. биол. наук. Л., 1954. 24 с.
Фалькенштейн Б. Ю. Некоторые эколого-географические закономерности динамики численности мышевидных грызунов // Защита растений. 1939. Т. 18. С. 121–134.
Фокин И. М. Тушканчики. Жизнь наших птиц и зверей . Л.: Изд-во Ленингр. ун-та, 1978. Вып. 2. 184 с.
Формозов А. Н. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930–1940 гг. // Материалы по грызунам. М.: Изд-во МОИП, 1948. Вып. 3. С. 3–110.
Хелевина С. А., Окулова Н. М. К экологии лесной мышовки (Sicista betulina) // Экология популяций: структура и динамика: Материалы всерос. совещ. М., 1988. С. 667–677.
Ходашева К. С. О географических особенностях структуры населения позвоночных животных // Зональные особенности населения позвоночных животных. М., 1966. 237 с.
Цветкова А. А. Особенности размножения лесной и степной мышовок на Южном Урале // Экология. 1978. № 3. С. 90–92.
Шенброт Г. И., Соколов В. Е., Гептнер В. Г., Ковальская Ю. М. Млекопитающие России и сопредельных регионов. Тушканчикообразные . М.: Наука, 1995. 576 с.
Штильмарк Ф. Р. Основные черты экологии мышевидных грызунов в кедровых лесах Западного Саяна // Фауна кедровых лесов Сибири и ее использование. М.: Наука, 1965. 359 с.
Юрлов К. Т., Юдин Б. С., Потапкина А. Ф. и др. К характеристике фауны мелких млекопитающих северной степи Барабинской низменности // Животный мир Барбы. Новосибирск: Наука, 1965. С. 184–207.
Godfrey G. K. Observation on the nature of the decline in numbers of two Microtus populations // J. Mammal. 1955. Vol. 36. № 2. P. 341–365.
Kubik I. Badania nad morrfologia I bioljgia smuzki (Sicista betulina Pall.) z Bialoweiskirgo Parku Narodowego // Ann. Univ. M. Cure-Skilod., Lublin, 1952. Vol. 11. № 1. S. 47–61.
Благодарности
Работа проводилась в плане выполнения Государственного задания Федерального исследовательского центра «Карельский научный центр РАН».

 © 2011 - 2026
© 2011 - 2026