



Введение
Температура окружающей среды является одним из важнейших абиотических факторов для пойкилотермных животных. Она определяет скорость химических реакций и является одним из ограничивающих условий роста и метаболизма животного. Для большинства пресноводных беспозвоночных диапазон существования может находиться в пределах от 0 °С до +40 °С, однако температурные границы активной жизнедеятельности намного уже (Проссер, 1977). Температура среды оказывает воздействие на функциональное состояние организма и процессы, его обуславливающие (Николаева, 2014; Балаева-Тихомирова, Кацнельсон, 2018; Голубев и др., 2017).
В меняющихся условиях среды сохранение жизнеспособности является наиболее насущной реакцией организма. Именно оборонительная, защитная реакция направлена на сохранение безопасности особи и выступает как доминирующая в отсутствие пищевых, половых и других стимулов (Сидоров, 2003).
Для пресноводных легочных моллюсков характерна защитная пассивно-оборонительная реакция в ответ на раздражающий стимул. Единственной оборонительной реакцией является сокращение щупальца и втягивание тела в раковину. Легкое прикосновение кисточкой или волоском к кожной поверхности вызывает локальное сокращение гладкой мускулатуры, более сильное – защитную реакцию, проявляющуюся в прикрытии тела моллюска раковиной (Осипов, 1982; Сидоров, 2003). У моллюсков самой чувствительной по отношению к температуре является нервная система. Тактильная стимуляция кожи разных участков, в том числе и щупальца, вызывает реакции в щупальцевом нерве моллюска в форме кратковременных вспышек импульсной активности. Действие внешнего раздражителя на моллюска в первую очередь отражается на его поведении, только через некоторое время происходят глубокие физико-химические изменения в его организме (Осипов, 1982; Гудимов, 2012). Поэтому поведенческие реакции при действии раздражающего стимула служат определенным индикатором для исследователя, а изучение оборонительной реакции позволяет быстро увидеть ответ на любое внешнее воздействие.
В оборонительном поведении моллюсков важную роль играют медиатор-специфичные системы нейронов, на эффективность работы которых влияют условия окружающей среды (Иерусалимский, 2009). Таким образом, температура среды может оказывать модулирующую роль на центральные нервные структуры моллюска, изменяя их активность, тем самым приводя к изменениям в ответных реакциях организма на предъявляемый стимул. В работе изучена зависимость оборонительного поведения от длительного воздействия пониженных температур.
Материалы
Работа выполнена на представителе пресноводных легочных моллюсков Helisoma duryi. Брюхоногие моллюски являются одним из удобных тест-объектов для изучения действия различных факторов внешней среды на состояние организма, т. к. они относительно быстро реагируют физиологическими, морфологическими, цитогенетическими и поведенческими изменениями (Толкачева, Хитров, 2015; Игнатов и др., 2013; Бондаренко, Боричева, 2004; Basopo, Ngabaza, 2015).
Маточную культуру моллюсков содержали в аквариумах объемом 5 литров с грунтом, высшей водной растительностью и системой принудительной аэрации и фильтрации при температуре +18–20 °С. В рацион питания моллюсков входили листья капусты, морковь и сухой хлопьевидный корм «Tetra min» для аквариумных рыбок в качестве витаминной добавки. Для изучения зависимости типов оборонительного поведения от размеров моллюсков использовали 232 особи. В эксперименте по изучению оборонительного поведения при воздействии температуры использовали моллюсков размерной категории от 7 до 10 мм, по 30 особей на вариант эксперимента. Моллюсков по одному помещали в отдельные лунки лабораторного планшета с отстоянной аэрированной водой и прикрывали крышкой, чтобы избежать возможность перемещения особей из одной лунки в другую. После адаптации моллюска к лунке планшета через 10 минут проводили воздействие нейлоновым градуированным волоском. Особи, подвергавшиеся температурному воздействию хотя бы в одном варианте эксперимента, в дальнейших исследованиях не использовались.
Методы
Промеры моллюсков проводили с использованием штангенциркуля, в частности измеряли высоту раковины. В зависимости от полученных данных моллюсков относили к одной из размерных категорий: 7.0–10.0 мм; 6–6.9 мм; 5.0–5.9 мм; 4.0–4.9 мм; 3.0–3.9 мм.
Изучение типов оборонительного поведения моллюсков проводили при температуре +18–20 ˚С. Моллюсков (n = 30, для каждого варианта) помещали в отдельные ячейки лабораторного планшета с отстоянной аэрированной водопроводной водой. После адаптации особей к условиям планшета в течение 10 минут начинали производить тактильное раздражение щупальца моллюска градуированным нейлоновым волоском с силой воздействия 0.01Н, 0.06Н и 0.001Н. Выделяли следующие типы ответов (по Сидорову, 2003): 1 – ретракция щупальца; 2 – прикрытие тела раковиной; 3 – игнорирование стимула. Определяли процент особей с разным типом реакций.
Для выбора экспериментальной температуры при изучении действия пониженных температур на оборонительное поведение проводили определение диапазона летальных температур при температуре акклимации +18–20 ˚С, период экспозиции 8 часов. Понижение температуры проводили с шагом в два градуса от +10 до -6 °С, экспозиция 8 часов. При необходимости шаг понижения температуры изменялся. Использовали три повторности, по 5 особей на повторность. Определение выживаемости моллюсков проводили по наличию двигательной активности. Испытания проводили с использованием климатической камеры (камера постоянной температуры и влажности, модель TH-ME-025). Время экспозиции и скорость понижения температуры в климатической камере задавали автоматически, скорость понижения температуры 0.1 ˚С в минуту до выбранного значения температуры.
Для изучения изменения оборонительного поведения при действии пониженных температур лабораторные планшеты с моллюсками (по 30 особей на вариант) помещали в климатическую камеру с заданными параметрами экспериментальных условий (температура, период экспозиции). После экспозиции отмечали характер реакции моллюска в ответ на тактильное раздражение щупальца нейлоновым волоском с силой воздействия 0.001Н. Аналогично проводили изучение реакции особей на данный стимул в контрольном варианте. Контрольной группой являлись моллюски, экспонировавшиеся в отстоянной аэрированной воде в лунках лабораторного планшета при температуре +18–20 ˚С. Повторное тактильное воздействие проводили через 20 минут после первого стимула.
Статистическая обработка данных проведена в программе Statistica 6.1. Статистическую значимость оценивали с помощью критерия соответствия χ2.
Результаты
В ходе исследования у моллюска Helisoma duryi отмечено три типа ответных реакций оборонительного поведения в ответ на тактильное раздражение нейлоновым градуированным волоском: игнорирование воздействия, ретракция щупальца, прикрытие тела раковиной (надвигание раковины).
Выявлено, что в зависимости от силы воздействия изменяется соотношение типов оборонительного поведения в ответ на раздражение. При силе воздействия 0.01Н реакция ретракции выявлена у 70 %, надвигание раковины у 20 %, игнорирование у 10 % особей. При силе воздействия 0.006Н 73.3 % отвечали ретракцией щупальца, 6.7 % надвиганием раковины, 20 % игнорированием стимула. При силе воздействия 0.001Н для 80 % особей отмечали реакцию ретракции, 3.4 % – надвигания, 36.6 % – игнорирования стимула (табл. 1).
Таблица 1. Изменение соотношения типов оборонительного поведения Helisoma duryi в зависимости от силы воздействия
| Тип ответной реакции | Сила воздействия, Н |
||
| 0.01 | 0.006 | 0.001 | |
| Число особей, шт. | |||
| Ретракция щупальца | 21 | 22 | 18 |
| Игнорирование | 3 | 6 | 11 |
| Прикрытие тела раковиной | 6 | 2 | 1 |
Получены статистически значимые отличия в соотношении типов оборонительного поведения при сравнении силы воздействия 0.01Н и 0.001Н (χ2 = 8.374, р < 0.05). В дальнейшем во всех вариантах эксперимента использовали нейлоновый волосок с силой воздействия 0.001Н.
На втором этапе изучали взаимосвязь соотношения типов оборонительных реакций и размера/возраста моллюска. В зависимости от размера раковины моллюски были разделены на группы: 1-я группа (69 особей) – 7.0–10 мм, 2-я группа (31 особь) – 6.0–6.9, 3-я группа (68 особей) – 5.0–5.9, 4-я группа (49 особей) – 4.0–4.9, 5-я группа (15 особей) – 3.0–3.9 мм.
Отмечено, что с увеличением размеров, а соответственно, и возраста моллюска соотношение типов оборонительных реакций меняется (рис. 1).
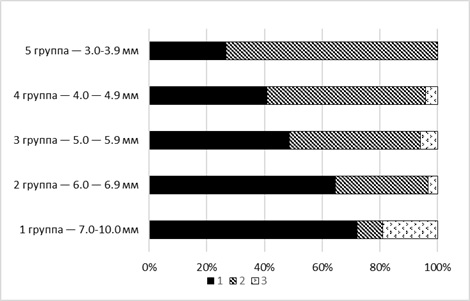
Рис. 1. Соотношение оборонительных реакций моллюсков Helisoma duryi в зависимости от их размера: 1 – ретракция щупальца, 2 – прикрытие тела раковиной, 3 – игнорирование воздействия
Fig. 1. The ratio of defensive reactions in Helisoma duryi mollusks depending on their size: 1 – tentacle retraction, 2 – covering the body with a shell, 3 – ignoring impact
Моллюски размером 3–3.9 мм проявляли только два варианта ответа: надвигание раковины (73.3 %) и ретракция щупальца (26.7 %). В группе от 4 до 4.9 мм реакцию игнорирования проявили 4.1 % особей, ретракцию щупальца – 40.8 %, надвигание раковины – 55.1 %. Процент моллюсков, отвечающих наиболее сильной реакций (надвиганием раковины в ответ на раздражение), уменьшается, а отвечающих ретракцией щупальца – возрастает с увеличением размеров особи (см. рис. 1). Связь между размером моллюска и проявлением типа оборонительной реакции носит статистически значимый характер (χ2 = 47.763, р < 0.01). На основании полученных данных для последующих экспериментов отбирали особей размером 7–10 мм.
Для изучения влияния температуры среды на поведение моллюсков возникла необходимость адекватного выбора температурного значения, позволяющего выявить воздействие пониженных температур, с одной стороны, и не приводящего к гибели особей при длительной экспозиции, с другой. В ходе исследования установлено, что в диапазоне температур от +10 ˚С до -2 ˚С при 8-часовой экспозиции выживали все особи. Гибель моллюсков наблюдали лишь при дальнейшем понижении температуры (табл. 2).
Таблица 2. Выживаемость моллюсков Helisoma duryi при воздействии пониженных температур
| Температура, t, °С | +10 | +8 | +6 | +4 | +2 | 0 | -2 | -4 | -4.5 | -5 | -6 |
| Выживаемость, % | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 80 | 63.2 | 40 | 0 |
Примечательным, на наш взгляд, является относительно высокая выживаемость моллюсков как при холодовых температурах, так и при морозных. Данный вид моллюска, обитающий в водоемах Северной Америки, является типичным представителем аквариумной малакофауны и интродуцирован в открытые природные и искусственные водоемы Украины (Сон, 2009).
С учетом полученных данных по диапазону нижних пессимальных температур для дальнейших исследований была выбрана температура +4 ˚С, являющаяся верхней границей холодового воздействия (Угаров, 2013) и не приводящая к гибели организма за выбранный период экспозиции.
Для изучения влияния пониженной температуры (+4 ˚С) на оборонительное поведение использовали варианты с экспозицией 60, 120, 240, 360 минут и 24 часа (сутки). Для каждого варианта экспозиции ставили отдельный контроль, тем самым получили серию вариантов для каждого временного промежутка.
При экспозиции в течение 60 минут при температуре +4 ˚С ответная реакция моллюсков на тактильное раздражение изменяется в сравнении с контролем. Несмотря на то что в ответных реакциях так же, как и в контрольном варианте, преобладает ретракция щупальца, отмечено увеличение количества особей моллюсков, отвечающих игнорированием тактильного воздействия. Так, 23.3 % особей в опытном варианте никак не отреагировали на раздражитель, тогда как в контроле таких особей выявлено 6.7 %. И только одна особь (3 %) отреагировала реакцией надвигания, в то время как в контроле число особей, реагирующих надвиганием раковины, составило 10 % (рис. 2). Однако полученные отличия не являются статистически значимыми (χ2 = 3.969, р < 0.05).
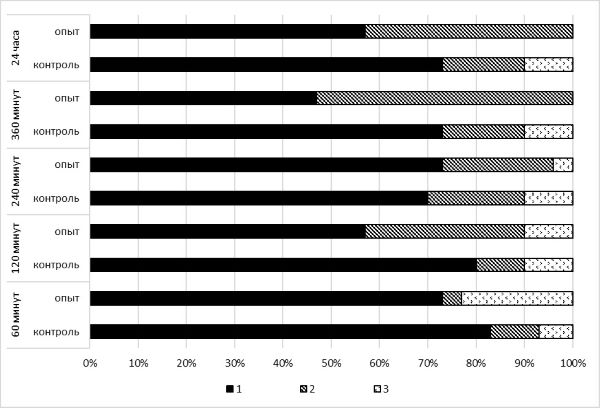
Рис. 2. Изменение соотношения типов поведения при температуре +4 ˚С в зависимости от времени экспозиции: 1 – ретракция щупальца, 2 – прикрытие тела раковиной, 3 – игнорирование воздействия
Fig. 2. Change in the ratio of defensive reactions of mollusks at a temperature of +4 ˚С, depending on the exposure time: 1 – tentacle retraction, 2 – covering the body with a shell, 3 – ignoring impact
Экспонирование моллюсков в течение 120 минут показало, что в ответных реакциях на тактильное раздражение по-прежнему преобладает реакция ретракции (57 %), но меньше, чем в контроле (80 %). В опытном варианте возрастает число моллюсков с сильной ответной реакцией на стимул – надвиганием раковины на передний конец тела (33 %). Тогда как в контроле число особей с реакцией надвигания составляет 10 %. Доля моллюсков, игнорирующих тактильное раздражение, остается такой же, как и в контрольной группе (3 %) (см. рис. 2). Выявленные отличия не являются статистически значимыми (χ2 = 4.964, р < 0.05).
При экспозиции моллюсков в течение 240 минут сохраняется преобладание ретракции как в контрольном (70 %), так и в опытном варианте (74 %). Доля сильных оборонительных реакций в опыте лишь на 3 % ниже, чем в контроле. Отмечено снижение числа особей, никак не отреагировавших на раздражение, в опытном варианте (3 %) по отношению к контролю (10 %) (см. рис. 2). Выявленные отличия не являются статистически значимыми (χ2 = 1.100, р < 0.05).
Несмотря на то что выявленные изменения в соотношении типов оборонительного поведения моллюсков при экспозиции от 60 до 240 минут статистически незначимы, однако наблюдается тенденция к уменьшению числа особей, игнорирующих стимул.
При экспозиции в течение 360 минут в опытном варианте отмечено отсутствие особей, игнорирующих раздражение. Выявлено только два типа оборонительных реакций – ретракция (47 %) и надвигание (53 %). В контрольном варианте – ретракция (73 %), надвигание (17 %), игнорирование (10 %). Изменение соотношения ответных реакций при холодовом воздействии (+4 °С) в течение 360 минут носит статистически значимый характер (χ2 = 10.540, р < 0.01).
Аналогичные изменения выявлены и при 24-часовой экспозиции. Воздействие пониженной температуры привело к проявлению только двух типов оборонительных реакций – ретракция (57 %) и надвигание (43 %). Моллюсков, отвечающих на раздражение игнорированием, не выявлено. Обнаруженные отличия носят статистически значимый характер по отношению к контролю (χ2 = 7.197, р < 0.05).
Таким образом, показано, что в зависимости от времени экспозиции моллюсков при пониженных температурах происходит изменение соотношений типов ответных оборонительных реакций. При наименьшем времени экспозиции (60 минут) отмечено наибольшее количество особей, не реагирующих на тактильное раздражение (снижение чувствительности – гипоэстезия). С увеличением времени экспозиции наблюдается постепенное увеличение количества особей, реагирующих на тактильное раздражение сильными оборонительными реакциями, т. е. отмечается усиление чувствительности особей (гиперэстезия). Через 360 минут экспозиции и более особей, игнорирующих раздражение, не выявлено.
Известно, что повторное предъявление стимула может приводить к развитию либо реакции привыкания, являющейся одной из форм физиологической адаптации животного к окружающей среде, либо сенситизации (облегчения), т. е. усилению ответной реакции (Осипов, 1982).
После повторного предъявления стимула через 20 минут в контрольном варианте статистически значимого изменения соотношений типов ответных реакций не выявлено (χ2 = 3.529, р < 0.05).
При повторном предъявлении стимула моллюскам, подвергавшимся холодовому воздействию (+4 ˚С), статистически значимые отличия в соотношении типов ответных реакций выявлены только после часовой экспозиции при +4 ˚С (χ2 = 13.233, р < 0.01). Сравнение проводили между первым и вторым предъявлением стимула для каждого варианта опыта (рис. 3).
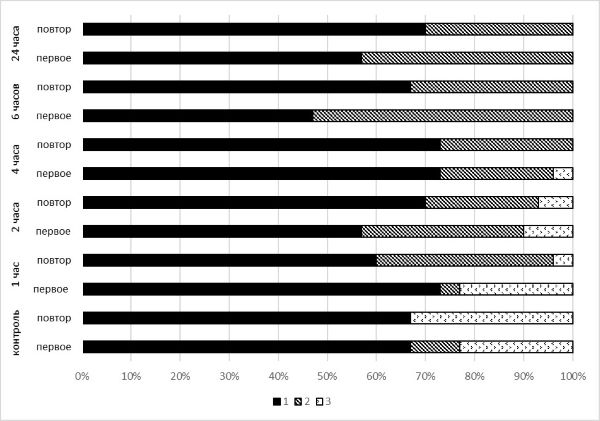
Рис. 3. Изменение соотношения ответных реакций оборонительного поведения моллюсков при повторном предъявлении стимула: 1 – ретракция щупальца, 2 – прикрытие тела раковиной, 3 – игнорирование воздействия
Fig. 3. Change in the ratio of defensive behavior responses in mollusks in repeated presentation of the stimulus: 1 – tentacle retraction, 2 – covering the body with a shell, 3 – ignoring impact
Повторное раздражение в других вариантах эксперимента не показало статистически значимых отличий в ответе на стимул.
Обсуждение
Оборонительное поведение, направленное на сохранение безопасности особи, преобладает над другими реакциями в отсутствие стимулов положительной эмоциональной направленности (пищевая, половая и др.) (Сидоров, 2003). В ответ на тактильное раздражение моллюски отвечают одним из типов оборонительных реакций: ретракцией (сокращение щупальца), отсутствием ответной реакции (игнорирование стимула), относящихся к проявлению слабых оборонительных реакций. Возможно полное прикрытие раковиной тела, или надвигание, которое относят к сильным оборонительным реакциям моллюска (Сидоров, 2003). Реакция надвигания является основной адаптивной реакцией, обеспечивающей эффективную защиту от неблагоприятных воздействий, и аналогична изолирующему рефлексу двустворчатых моллюсков – реакции закрытия створок (Гудимов, 2012).
В зависимости от силы воздействия раздражителя соотношение типов оборонительных реакций может изменяться. Наиболее частым вариантом ответа моллюска на стимул при всех трех вариантах силы воздействия (от 0.001Н до 0.01Н) является сокращение щупальца (ретракция). Однако необходимо отметить, что изменяется соотношение реакций игнорирование стимула и надвигание раковины. Чем больше сила воздействия, тем чаще проявляются сильные оборонительные реакции организма, в частности надвигание раковины на передний конец, полное прикрытие тела. По закону силы величина ответа зависит от силы раздражителя, т. е. чем больше сила раздражителя, тем больше величина ответа (Ноздрачев и др., 1991). По мере снижения силы воздействия уменьшается процент особей, реагирующих на стимул надвиганием раковины (сильная оборонительная реакция) и увеличивается процент особей, отвечающих на тактильный стимул игнорированием (слабая оборонительная реакция).
Изучение зависимости соотношения типов оборонительных реакций от размера моллюска показало, что чем меньше размер моллюска, а соответственно, и его возраст, тем больший процент особей отвечает на раздражение щупальца надвиганием раковины вплоть до полного прикрытия тела. При этом минимизируется количество особей, не реагирующих на воздействие (реакция игнорирования) и снижается количество моллюсков, реагирующих ретракцией щупальца. Так, у группы моллюсков от 3.0 до 3.9 мм 73 % особей ответили на стимул надвиганием раковины, лишь 27 % особей – ретракцией щупальца, а особей, никак не отреагировавших на воздействие, выявлено не было. Такая ответная реакция моллюсков наименьших размеров может быть связана как с большей нагрузкой на единицу площади соприкосновения при равной силе воздействия, так и с физиологическими процессами.
Информация о тактильном раздражении от первично сенсорных нейронов поступает к мотонейронам и параллельно ко многим медиатор-специфичным нейронным системам, в т. ч. к премоторным интернейронам, способным запускать оборонительные реакции. У ряда животных описаны «командные» интернейроны, структурирующие сенсорную информацию, необходимые и достаточные для запуска определенного поведенческого акта (Балабан, Коршунова, 2011). Известно, что командные нейроны оборонительного поведения брюхоногого моллюска Helix, расположенные симметрично в левых и правых париетальных ганглиях, выполняют сходные функции – запускают оборонительное поведение в ответ на опасные стимулы и являются главным пластическим звеном условных оборонительных рефлексов (Гринкевич, Зачепило, 2017).
Важную роль в оборонительном поведении виноградной улитки Helix играют серотонин и нейропептид FMRFамид, т. к. серотонин содержится в модуляторных нейронах, получающих разнообразную сенсорную информацию об окружающей среде, а FMRFамид – в командных нейронах оборонительного поведения, отвечающих за оборонительные реакции, включая втягивание всего тела в раковину. Помимо этого в регуляции оборонительного поведения имеются CNP- и ГАМК- нейропептиды. Наличие системы модуляторных нейронов позволяет отвечать на значимые воздействия внешней среды как единое целое (Асеев, 2005; Балабан, Коршунова, 2011; Иерусалимский, Балабан, 2010). Оборонительное поведение может отличаться в зависимости от возраста моллюсков. Предполагается, что у новорожденных особей Pulmonata сформировано большинство клеточных элементов, отвечающих за необходимые физиологические процессы, однако процессы нейрогенеза, в основном модуляторных нейронов и других опосредующих клеток в онтогенезе продолжаются и могут лежать в основе меняющегося с возрастом поведения животного. Показано, что у ювенильных особей Helix слабо развита серотонинергическая система нейронов, играющих модулирующую роль в оборонительном поведении, однако отмечается наличие нейронов, содержащих ГАМК, но отсутствующих впоследствии у взрослых животных (Иерусалимский, 2009; Иерусалимский, Балабан, 2010).
Таким образом, возможно, что ответная реакция моллюска на тактильное воздействие на щупальце обусловлена синаптической передачей сигнала в командных нейронах оборонительного поведения, а тип реакции (ретракция щупальца, надвигание раковины, игнорирование) – работой медиатор-специфичных систем нейронов, модулирующих оборонительное поведение. Поскольку развитие медиатор-специфичных систем имеет возрастные особенности, это может также являться причиной различий в соотношении ответных реакций моллюсков в зависимости от их размеров.
Анализ изменений в соотношении типов оборонительного поведения при воздействии пониженных температур показал, что статистически значимые отличия в изменении соотношения типов оборонительного поведения происходят только при 6- и 24-часовой экспозиции моллюсков при температуре +4 °С. При данных экспозициях изменения в оборонительном поведении заключаются в отсутствии особей, отвечающих на воздействие игнорированием. Все опытные особи отвечают либо ретракцией щупальца, либо надвиганием раковины. Причем увеличивается доля особей, проявляющих наиболее сильную оборонительную реакцию (надвигание раковины), за счет уменьшения числа особей, отвечающих ретракцией. Необходимо отметить, что рост числа особей, реагирующих на стимул надвиганием раковины, за счет снижения числа особей, игнорирующих воздействие, наблюдается по мере увеличения времени периода экспозиции.
Возможно, такая реакция моллюсков связана с влиянием температуры на соответствующие нейроны, контролирующие оборонительное поведение. Показано, что в ЦНС моллюсков между нейронами оборонительных, модуляторных, нейросекреторных клеток имеются электротонические контакты. Электрические контакты чувствительны к действию высоких температур, а химические – к действию низких (Сидоров, 2012).
Известно, что нейропептиды отличаются по степени зависимости от воздействий внешней среды. Их пластичность обусловлена как типом нейропептида, так и местом локализации. Экспрессия нейропептидов семейства CNP зависит от внешних воздействий, в то время как экспрессия FMRFамида и педального пептида не зависит (Иерусалимский, 2009). Нейропептиды, играющие роль нейромодуляторов, могут влиять на выброс медиатора из пресинаптической терминали и менять временные и амплитудные параметры постсинаптического ответа (Асеев, 2005). Таким образом, можно предположить, что по мере увеличения времени холодового воздействия изменяется экспрессия нейропептидов разных групп, а соответственно, и амплитудные параметры постсинаптического ответа в сторону усиления ответной реакции организма на стимул.
При рассмотрении полученных результатов с точки зрения теории адаптации можно допустить, что реакция моллюска на холодовое воздействие при экспонировании от 60 минут до 24 часов отражает разные стадии адаптационного процесса организма. При экспозиции от 60 до 240 минут проявляется неспецифическая реакция на воздействие, отражающая начальную реакцию систем организма. При данных экспозициях изменения в оборонительном поведении проявляются, но не носят статистически значимый характер. После 6-часовой экспозиции происходит переход организма к новому состоянию временной устойчивости функциональных систем, заключающемуся в изменениях проявления типов оборонительного поведения. Переход организма на эту «новую» стадию устойчивости систем подтверждается аналогичными изменениями в типах оборонительных реакций и после 24-часового воздействия.
При повторном предъявлении стимула через 20 минут после первого раздражения щупальца также отмечено изменение соотношения типов ответных реакций. В контрольном варианте при повторном предъявлении стимула отмечается уменьшение доли особей, реагирующих надвиганием, за счет увеличения доли особей, отвечающих на раздражитель игнорированием. Это отражает типичную модификацию поведения, выражающуюся в понижении амплитуды ответа организма на повторное предъявление стимула. Таким образом, в нормальных условиях появляются особи, отвечающие привыканием (габитуацией) в ответ на повторное воздействие.
Повторное воздействие на моллюсков, экспонировавшихся в течение 60 минут при +4 °С, показало противоположный результат – увеличение доли особей, реагирующих надвиганием раковины за счет уменьшения количества особей, отвечающих игнорированием. Такая реакция сенситизации (облегчения) может быть обусловлена повышением амплитуды, частоты или продолжительности ответа на повторное предъявление стимула. Возможно, это связано с первоначальной активацией серотонинергических модуляторных нейронов при опасных для животных параметрах окружающей среды (Балабан, Коршунова, 2011).
В других вариантах опыта (2, 4, 6, 24 часа) статистически значимых изменений в соотношении типов оборонительного поведения после повторного предъявления стимула не выявлено. На наш взгляд, это может свидетельствовать о постепенном привыкании организма к новым условиям среды и его переходу на новую стадию устойчивости функциональных систем. Причем отсутствие изменений в поведенческих реакциях на повторный стимул через 20 минут после первого воздействия свидетельствует о достаточно глубоких перестройках в организме и возможном преобладании деятельности одной из модулирующих систем нейропептидов. С учетом того что FMRFамид играет существенную роль в формировании привыкания, оказывая тормозное действие на поведенческие реакции в ответ на стимулы, и его экспрессия не зависит от внешних воздействий (Гринкевич, Зачепило, 2018; Иерусалимский, 2009), вероятно, в условиях пониженных температур именно данные нейропептиды играют ключевую роль в ответной реакции на повторный стимул.
Заключение
- Характеристики оборонительного поведения моллюсков Helisoma duryi зависят от силы оказываемого воздействия. По мере снижения силы воздействия уменьшается процент особей, реагирующих на стимул надвиганием раковины (сильная оборонительная реакция), и увеличивается процент особей, отвечающих на тактильный стимул игнорированием (слабая оборонительная реакция).
- Выявлена взаимосвязь между размером моллюска и типом оборонительного поведения. С уменьшением размеров моллюсков увеличивается доля особей, отвечающих надвиганием раковины, за счет уменьшения числа особей, отвечающих ретракцией щупальца и/или игнорирующих стимул.
- Холодовое воздействие (+4 °С) в зависимости от времени экспозиции приводит к изменению типов оборонительного поведения. Статистически значимые отличия в изменении соотношения типов оборонительного поведения выявлены при 6- и 24-часовой экспозиции моллюсков, они выражены в увеличении доли особей, реагирующих надвиганием раковины; снижении числа особей, отвечающих ретракцией щупальца; отсутствии особей, игнорирующих стимул.
- Повторное предъявление стимула приводит к разнонаправленным изменениям в оборонительном поведении в зависимости от периода холодового воздействия. Кратковременная экспозиция в течение 60 минут приводит к сенситизации ответных реакций – увеличивается доля особей, отвечающих на стимул надвиганием раковины. При увеличении времени экспозиции повторное предъявление стимула не вызывает статистически значимых изменений в соотношении типов оборонительного поведения по отношению к первому воздействию.
Библиография
Асеев Н. А. Обнаружение и функциональное исследование нейропептидов, гомологичных пептидам CNP улитки Helix lucorum L., в нервной системе других беспозвоночных : Автореф. дис. … канд. биол. наук. М., 2005. 24 с.
Балабан П. М., Коршунова Т. А. Сетевые, клеточные и молекулярные механизмы пластичности в простых нервных системах // Успехи физиологических наук. 2011. Т. 42. № 4. С. 3–19.
Балаева-Тихомирова О. М., Кацнельсон Е. И. Особенности обмена веществ Planorbarius corneus в зависимости от sсезона года и местообитания // Веснік ВДУ. 2018. № 1. С. 66–74.
Бондаренко В. Ф., Боричева Е. С. Изменение эффективности отдельных сахаров в стимуляции пищевого поведения у пресноводных легочных моллюсков прудовика обыкновенного (Lymnaea stagnalis) и катушки роговой (Planorbarius corneus L.) в зависимости от пищевого опыта // Сенсорные системы. 2004. Т. 18. № 1. С. 79–82.
Голубев А. П., Хомич А. М., Долматова В. В., Толкачева Т. А. Действие гипертермии разной продолжительности на модельные тест-организмы // Веснік Віцебскага дзяржаўнага ўніверсітэта. 2017. № 4 (97). С. 34–39.
Гринкевич Л. Н., Зачепило Т. Г. Регуляция ацетилирования гистона Н4 в командных нейронах оборонительного поведения моллюска Helix // Рецепторы и внутриклеточная сигнализация: Сборник статей Международной конференции. СПб., 2017. С. 222–227.
Гринкевич Л. Н., Зачепило Т. Г. Регуляция ацетилирования гистона Н4 в центральной нервной системе и командных нейронах оборонительного поведения моллюска Helix серотонином и нейропептидом FMRFамидом // Вавиловский журнал генетики и селекции. 2018. Т. 22. № 5. С. 606–610.
Гудимов А. В. Первые записи поведенческих реакций двустворчатых моллюсков исландского гребешка Chlamys islandica и модиолуса Modiolus modiolus // Вестник Камчатского государственного технического университета. 2012. № 20. С. 50–55.
Игнатов Ю. Д., Тюренков И. Н., Вислобоков А. И., Перфилова В. Н., Мокроусов И. С. Изменения трансмембранных ионных токов нейронов моллюсков под влиянием соединения РГПУ-207 // Биологические мембраны. 2013. Т. 30. № 3. С. 206–213.
Иерусалимский В. Н. Функциональная регуляция и онтогенез медиатор-специфичных систем нейронов беспозвоночных : Автореф. дис. … д-ра биол. наук. М., 2009. 39 с.
Иерусалимский В. Н., Балабан П. М. Серотонинергическая система нейронов в ЦНС виноградной улитки: морфология, онтогенез, контроль поведения // Журнал высшей нервной деятельности. 2010. Т. 60. № 5. С. 515–524.
Николаева Н. Е. Влияние абиотических факторов на изменение фототропизма у брюхоногих моллюсков Lymnaea lagotis Schrank и Segmentina montgazoniana Borguignat // Вестник ТвГУ. Серия «Биология и экология». 2014. № 3. С. 42–52.
Ноздрачев А. Д., Баранникова И. А., Батуев А. С. Общий курс физиологии человека и животных . Кн. 1: Физиология нервной, мышечной и сенсорной систем. М.: Высшая школа, 1991. 512 с.
Проссер Л. Сравнительная физиология животных . М.: Мир, 1977. Т. 2. 571 с.
Сидоров А. В. Влияние температуры на легочное дыхание, оборонительные реакции и локомоторное поведение пресноводного легочного моллюска Lymnaea stagnalis // Журнал высшей нервной деятельности им И. П. Павлова. 2003. Т. 53. № 4. С. 513–517.
Сидоров А. В. Эволюция межклеточной коммуникации и структурная организация мозга // Журнал эволюционной биохимии и физиологии. 2012. Т. 48. № 4. С. 323–328.
Сон М. О. Моллюски-вселенцы на территории Украины: источники и направления инвазии // Российский журнал биологических инвазий. 2009. № 2. С. 37–48.
Толкачева Т. А., Хитров Д. М. Изучение физиологических, биохимических показателей тканей пресноводных моллюсков для экологического мониторинга водоемов // Наука – образованию, производству, экономике: Материалы XX (67) Регион. науч.-практ. конф. преподавателей, науч. сотрудников и аспирантов. Изд-во ВГБУ им. П. М. Машерова, 2015. С. 79–81.
Угаров Г. С. Теоретические основы гипобиологии // Фундаментальные исследования. 2013. № 10. С. 1280–1283.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда . Книга 1 / Пер. с англ. М. Д. Гроздовой, Г. И. Рожковой; Под ред Е. М. Крепса. М.: Мир, 1982. 426 с.
Basopo N., Ngabaza T. Toxicological Effects of Chlorpyrifos and Lead on the Aquatic Snail Helisoma duryi // Advances in Biological Chemistry. 2015. № 5. Р. 225–233.

 © 2011 - 2026
© 2011 - 2026