



Введение
Клен ясенелистный (Acer negundo L.) является крайне агрессивным инвазионным видом. Во вторичном ареале он способен обитать в широком диапазоне условий, слабо поражается болезнями и вредителями, может быстро образовывать многоярусные заросли. Клен ясенелистный нередко вытесняет аборигенные виды деревьев, создавая тем самым угрозу биологическому разнообразию (Виноградова и др., 2010; Marozas et al., 2015; Straigyte et al., 2015). Поэтому его проникновение крайне нежелательно для особо охраняемых природных территорий (ООПТ) (Арбузова, 2005; Braun et al., 2016; Dumitraşcu, Bălteanu, 2014).
Для ряда региональных и федеральных ООПТ в степной и лесостепной зоне отмечено внедрение клена ясенелистного (Арбузова, 2005; Емельянов, Фролова, 2011; Рыжков, Рыжкова, 2012; Агишев, 2017; Фатьянова и др., 2017; Саксонов, 2018). В их число входит участок Ямская степь заповедника «Белогорье». В его охранной зоне образовалась заросль клена ясенелистного, примыкающая к границе заповедника (Арбузова, Украинский, 2017).
В большинстве существующих исследований анализируется только текущее распространение клена ясенелистного (Camenen et al., 2016). Временная динамика процесса инвазии исследуется достаточно редко. Существующие исследования такого рода сосредоточены в основном на оценке изменения площади, занятой зарослями клена ясенелистного (Колтунова, Кузьмин, 2017). Представленная работа направлена на восстановлении истории расселения клена ясенелистного в охранной зоне Ямской степи. Эта задача включает в себя определение численности и плотности размещения деревьев в разные хронологические срезы, определение типа пространственного распределения, выявление основных направлений расселения.
Материалы
Исследуемая территория находится в охранной зоне участка Ямская степь заповедника «Белогорье» (Белгородская область, Губкинский городской округ). Граница исследуемой территории проведена по периметру заросли клена ясенелистного. Эта заросль имеет форму полосы, протянувшейся вдоль межевой канавы, разделяющей второй квартал Ямской степи и охранную зону заповедника. С севера на юг эта полоса протягивается на 177 м. Ширина полосы колеблется от 13 до 17 м. Площадь исследуемой территории составляет 3296 м2.
С запада к исследуемой территории, в центральной ее части, примыкает поселок Загорный. К настоящему времени он является фактически нежилым, за исключением кордона заповедника (рис. 1). Большая часть строений находится в полуразрушенном состоянии. На данный момент поселок уже можно считать постселитебной территорией. В Белгородской области для таких территорий характерно формирование лесных сообществ, в которых клен ясенелистный выступает видом-средообразователем (Голеусов, Артищев, 2018).
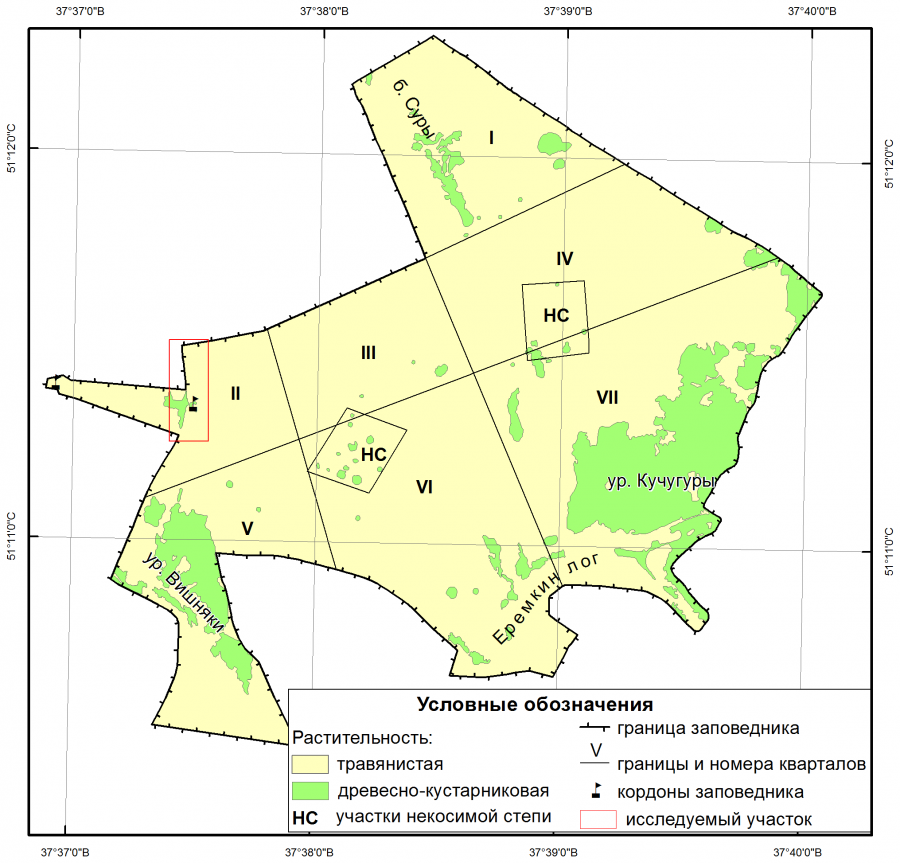
Рис. 1. Расположение исследуемой территории
Fig. 1. Location of the study area
На территории поселка Загорный сохранилась рядовая посадка, отделявшая усадьбы поселка от огородов. Эта посадка является предполагаемым и наиболее вероятным источником заноса клена ясенелистного в охранную зону Ямской степи. В результате заноса из поселка в межевой канаве и вдоль нее сформировалась полночленная популяция клена ясенелистного, размножение в которой происходило как семенным, так и вегетативным путем. На территорию прилегающего второго квартала заповедника клен ясенелистный практически не проник. Здесь было обнаружено только одно дерево (Арбузова и др., 2014). Расселению клена ясенелистного внутри заповедного участка мешал режим периодического сенокошения, поддерживаемый в Ямской степи, и наличие плотной степной дернины (Арбузова, Украинский, 2017).
Исходные данные собирались осенью 2014 и 2015 гг. В это время проводилась расчистка межевой канавы и прилегающей к ней полосы охранной зоны от клена ясенелистного. Параллельно с этим выполнялось картографирование расположения спиленных деревьев (Арбузова, Украинский, 2017). Их положение фиксировалось относительно межевой канавы. Для этого измерялось расстояние от дерева до межевой канавы и расстояние, пройденное по межевой канаве от начальной точки. По этим измерениям на миллиметровой бумаге была составлена первичная схема размещения деревьев. Географическая привязка отсканированной схемы и оцифровка положения деревьев выполнялись в программе ArcGIS.
Чтобы восстановить историю распространения клена ясенелистного, необходимо было проанализировать возраст деревьев. Для этого по спилам было посчитано число годовых колец. Кроме этого измерялся диаметр ствола. Так как сечение ствола имело не круглую, а эллиптическую форму, то измерялось два диаметра: больший и меньший. У части деревьев от одного корня вырастали два ствола и более. В таких случаях для анализа брались измерения, выполненные по самому старому стволу.
Методы
Главной проблемой при определении возраста деревьев была частичная неполнота данных, т. к. не всегда можно было подсчитать точное число годовых колец. Среди деревьев присутствовали экземпляры с выгнившей сердцевиной. Диаметр выгнившей сердцевины соответствовал приблизительно 2–3 годам возраста дерева. Также среди деревьев были мертвые экземпляры с полностью прогнившим, трухлявым стволом. Для них возраст был восстановлен с помощью линейной регрессии. Была создана регрессионная модель, связывающая число годовых колец на спиле с большим и малым диаметром спила. Для построения модели использована выборка из деревьев с точным подсчитанным числом годовых колец. Все расчеты проведены в среде статистических вычислений R (R Core Team, 2020).
Ретроспективный анализ распространения клена ясенелистного выполнялся по семи временным срезам, которые были взяты с интервалом в пять лет, от 2015 до 1985 г. Изучение распределения деревьев в пространстве выполнено в среде статистических вычислений R с использованием дополнительных пакетов spatstat (Baddeley et al., 2015; Bivand et al., 2013) и dixon (Dixon, 2002).
Для визуализации положения деревьев в каждом временном срезе был построен растр локальной плотности. При его создании использован переменный радиус поиска, определяемый по методу Абрамсона (Abramson, 1982). Кроме визуального анализа, расположение деревьев оценивалось количественными методами. С помощью теста Кларка – Эванса определялся тип пространственного распределения (Clark, Evans, 1954). С помощью теста Диксона (Dixon, 2002) оценивалась степень сегрегации молодых деревьев, появившихся за последние пять лет, и старых деревьев.
Результаты
На исследуемой территории было закартографировано положение 298 деревьев клена ясенелистного. У 220 деревьев точно подсчитано число годовых колец. У 53 деревьев из-за выгнившей сердцевины число годовых колец подсчитано частично (возраст недооценен на 2–3 года). У 25 деревьев ствол прогнил полностью, и подсчитать число годовых колец оказалось невозможным. Для этих деревьев возраст был восстановлен на основе связей между числом годовых колец и диаметрами спила (малым и большим). Для выборки деревьев, у которых полностью подсчитано число годовых колец, отмечается высокая теснота этих связей. Коэффициент корреляции Спирмена для числа колец и малого диаметра составляет ρ = 0.81 при p = 1.49·10-52. Коэффициент корреляции Спирмена для числа колец и большого диаметра составляет ρ = 0.81 при p = 3.07·10-51. Число годовых колец для полностью прогнивших стволов было рассчитано по следующей регрессионной модели:
C = 2.31 + 0.64 · D1 – 0.01 · D1 · D2 + 0.39 · D2,
где C – число колец на спиле, D1 – малый диаметр спила, D2 – большой диаметр спила. Модель, использованная для восстановления возраста деревьев с трухлявыми стволами, является статистически значимой (по результатам теста Вальда F = 128.47, p = 8.85 · 10-48) и имеет коэффициент детерминации R2 = 0.64. Среднеквадратическая ошибка модели составляет 3.87 года.
С учетом восстановленных значений возраст деревьев клена ясенелистного колеблется от 3 до 35 лет. Средний возраст составляет 13.3 года, а стандартное отклонение возраста – 6.16 года. Распределение по пятилетним возрастным группам неравномерное, что подтверждается статистически (по результатам теста хи-квадрат χ2 = 230.52, p = 5.92 · 10-47). Наблюдается смещение числа деревьев в пятилетних возрастных группах в сторону молодых возрастов (рис. 2).
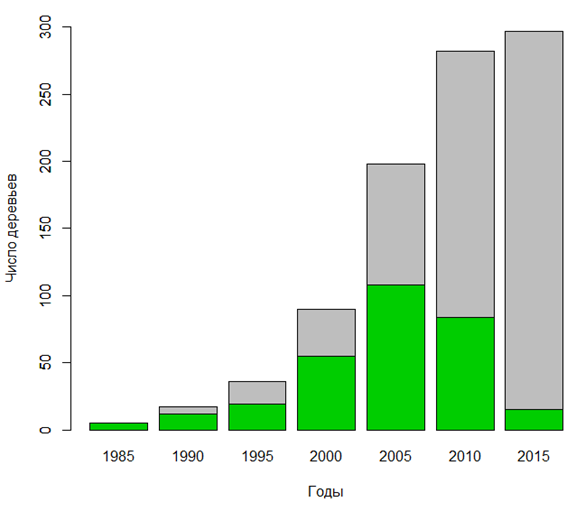
Рис. 2. Численность клена ясенелистного по пятилетним возрастным группам (зеленый цвет – численность деревьев, появившихся за прошедшие 5 лет, серый цвет – накопленная численность с начала распространения)
Fig. 2. The number of boxelder maple by five-year age groups (green color – abundance over the past 5 years, gray color – accumulated abundance from the beginning of distribution)
Интенсивность увеличения количества деревьев в 1985–2015 гг. была непостоянной. До 1995 г. численность клена ясенелистного увеличивалась слабо. В среднем за год появлялось 3.1 нового дерева. В 1995–2010 гг. интенсивность увеличения количества деревьев резко выросла. В этот период за год в среднем появлялось 16.4 нового дерева. После 2010 г. интенсивность прироста численности резко снизилась, составляя 1.5 нового дерева в год. Период усиливающейся интенсивности появления новых деревьев (с 1985 по 2005 г.) совпадает со временем, когда клен ясенелистный заселял полосу вдоль межевой канавы, которая имеет лучшие условия увлажнения (рис. 3). После того как в этой полосе образовалась сомкнутая заросль и клен ясенелистный стал расселяться (в период 2005–2015 гг.) уже на прилегающие территории с худшими условиями увлажнения, интенсивность появления новых деревьев начинала снижаться.
Возраст наиболее старых деревьев, составляющий 35 лет, указывает на 1980 г. как время начала расселения клена ясенелистного вдоль межевой канавы. Большая часть исследуемой территории находится в пределах радиуса 50 м от поселка Загорный. Как раз на такое расстояние разносятся ветром крылатки клена ясенелистного (Sachse et al., 1991). Однако первоначальное заселение произошло не по всей длине межевой канавы, а преимущественно в ее южной части (см. рис. 3). Вероятно, первые деревья закрепились на участке с наиболее благоприятными условиями произрастания. В условиях Ямской степи главным лимитирующим фактором является увлажнение. Межевая канава второго квартала Ямской степи идет с севера на юг по склону вниз, от водораздела до балки Вишняки. Соответственно с севера на юг нарастает увлажнение. И первые деревья закрепились в южной части межевой канавы на расстоянии максимального ветрового переноса крылаток. Примечательно, что после того как межевая канава поворачивает на восток, вдоль северной границы второго квартала, клен ясенелистный в ней перестает обнаруживаться. В 2014 г. там не обнаружено ни одного дерева клена ясенелистного, хотя другие виды деревьев встречаются (Арбузова и др., 2014). В этой части заповедника межевая канава заходит на водораздел и уходит за пределы 50-метрового радиуса от поселка Загорный.
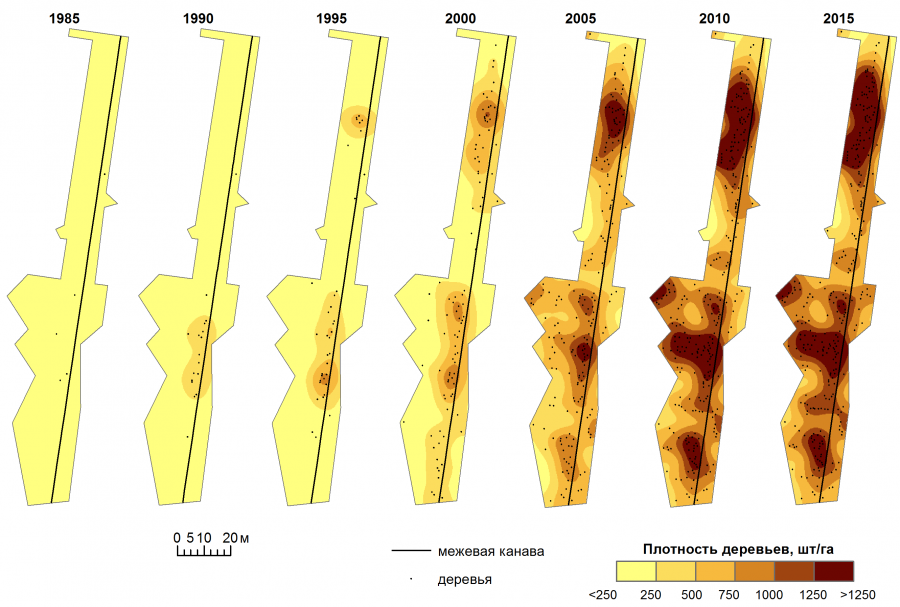
Рис. 3. Распределение деревьев клена ясенелистного на исследуемой территории в различные годы
Fig. 3. Distribution of boxelder maple in the study area in different years
На открытых местах с плодородными почвами, чему в полной мере соответствует охранная зона Ямской степи, клен ясенелистный начинает плодоносить с 5 лет (Medrzycki, 2002). Поэтому к 1995 г. он мог заноситься не только из поселка Загорный, но и с деревьев, поселившихся у межевой канавы.
До 1990 г. новые деревья появлялись только в первом очаге расселения. К 1995 г. формируется второй очаг расселения – клен ясенелистный закрепляется в северной части исследуемого участка. Во всех последующих временных срезах (2000–2015 гг.) этим двум первичным очагам расселения будут соответствовать зоны максимальной плотности распространения клена ясенелистного. К 2015 г. формируется специфическая картина распределения плотности, когда при перемещении вдоль межевой канавы регулярно чередуются участки повышенной и пониженной плотности деревьев. При этом протяженность таких участков приблизительно одинакова. Подобная картина наблюдается при распространении клена ясенелистного в поймах рек (Абрамова и др., 2019; Valantinaite et al., 2011) и вдоль автомобильных дорог (Леонтьев, Зверева, 2016).
Ритмичная смена участков с повышенной и пониженной плотностью деревьев вдоль линейных объектов (как естественных, так и антропогенных) обусловливается сочетанием размеров и формы территории с расстоянием ветрового переноса. В случае речных долин, автомобильных дорог и межевых канав территория расселения представляет собой узкую, но протяженную полосу с благоприятными для расселения условиями. При этом длина полосы кратно больше, чем расстояние ветрового переноса семян. Мы предполагаем, что аналогично искусственным рвам и канавам расселение клена ясенелистного может происходить вдоль промоин на эродированных склонах.
На примере Ямской степи видно, что достаточно, чтобы протяженность участка превышала не максимальное, а среднее расстояние ветрового переноса (для различных видов рода Acer среднее расстояние ветрового переноса в 3–4 раза меньше максимального) (Евстигнеев и др., 2017). В таком случае вокруг первых заселившихся деревьев формируются участки с повышенной плотностью клена ясенелистного. Но чередование участков высокой и низкой плотности не всегда может быть таким четким, как на исследуемой нами территории. Вдоль западной границы второго квартала Ямской степи присутствовал один источник заноса клена ясенелистного. Наличие же нескольких источников заноса может существенно усложнить картину изменения плотности. Помимо ветрового переноса, для клена ясенелистного отмечается перенос водными потоками (Акатов и др., 2014). Этот дополнительный путь переноса может усложнять картину пространственного распределения плотности клена ясенелистного, расселяющегося в поймах рек.
До 2000 г. две первые заросли клена ясенелистного (северная и южная), появившиеся вокруг первичных очагов распространения, продолжают разрастаться вдоль межевой канавы. Одновременно с этим начинается распространение в стороны от межевой канавы. В 2000 г. деревья клена ясенелистного появляются в самой южной части исследуемого участка, удаленной от поселка Загорный более чем на 50 м. Сюда новые деревья заселились уже явно из полосы вдоль межевой канавы.
К 2005 г. северная и южная заросль клена ясенелистного соединяются. И после 2005 г. основное расселения идет уже не вдоль межевой канавы, а вбок от нее. Формируется сплошная сомкнутая заросль.
Кроме увеличения плотности размещения и расширения границы распространения клена ясенелистного в период 1985–2015 гг. менялся тип пространственного распределения деревьев. Для временных срезов 1985 и 1990 гг. величина критерия Кларка – Эванса статистически значимо не отличается от 1.0 (табл. 1). Это указывает на случайное пространственное распределение. А с 1995 по 2015 г. отмечается групповое пространственное распределение, когда значения критерия Кларка – Эванса статистически значимо меньшие, чем 1.0 (см. табл. 1).
Степень выраженности группового распределения в пространстве изменялась, что видно по динамике значения критерия Кларка – Эванса. Уменьшение этого значения говорит об усилении тенденции к группированию, а увеличение – об ослаблении этой тенденции.
В 1995 г. отмечается наибольшая выраженность группового распределения. Это совпадает со временем существования двух четко разделенных групп деревьев – первых двух очагов распространения клена ясенелистного вдоль межевой канавы. По мере срастания зарослей, формирующихся вокруг двух очагов первичного расселения, выраженность группового распределения ослабевает, а величина критерия Кларка – Эванса растет.
Таблица 1. Результаты теста Кларка – Эванса
| Год | R критерий | р значение |
| 1985 | 1.32 | 0.17 |
| 1990 | 0.8 | 0.12 |
| 1995 | 0.54 | 1.09·10-7 |
| 2000 | 0.84 | 0.004 |
| 2005 | 0.91 | 0.02 |
| 2010 | 0.92 | 0.01 |
| 2015 | 0.93 | 0.03 |
Новые деревья клена ясенелистного появлялись как за пределами формирующейся заросли, так и внутри ее. В первом случае расширялись границы заросли, а во втором – увеличивалась плотность размещения деревьев. В разные периоды жизни заросли соотношение этих двух процессов отличалось. О том, какой из них преобладал, можно судить по степени пространственной сегрегации молодых (младше 5 лет) и старых деревьев (табл. 2).
Таблица 2. Результаты теста Диксона
| Год | Сопоставляемые возрастные группы | C критерий | р значение |
| 1990 | Младше 5 лет и 5–10 лет | 1.88 | 0.39 |
| 1995 | Младше 5 лет и 5–15 лет | 4.3 | 0.12 |
| 2000 | Младше 5 лет и 5–20 лет | 16 | 0.0003 |
| 2005 | Младше 5 лет и 5–25 лет | 11.52 | 0.003 |
| 2010 | Младше 5 лет и 5–30 лет | 1.85 | 0.4 |
| 2015 | Младше 5 лет и 5–35 лет | 27.37 | 1.14 · 10-6 |
Для временных срезов 1990 и 1995 гг. молодые и старые деревья территориально не отделены друг от друга. Новые деревья появляются в основном в пределах первого очага расселения, расположенного в южной части исследуемой территории. В следующие два временных среза (2000 и 2005 гг.) новые деревья появляются в основном за пределами заросли. По мере срастания двух зарослей, сформировавшихся вокруг первых очагов расселения, величина С-критерия, показывающего степень сегрегации, уменьшается. Резко она увеличивается в 2015 г., достигая максимального значения за весь анализируемый период времени. Возможно, в это время начинался следующий этап расширения заросли клена ясенелистного, который был искусственно прерван.
Изменение степени сегрегации молодых (младше пяти лет) и старых деревьев (см. табл. 2) связано с влиянием древесного полога на прорастание и развитие клена ясенелистного. С одной стороны, известно, что клен ясенелистный теневынослив и хорошо прорастает под пологом деревьев (Костина и др., 2013). Проростки и молодые особи всегда встречаются внутри зарослей клена ясенелистного (Дайнеко и др., 2017). При этом затенение все-таки оказывает влияние на развитие проростков под пологом деревьев (Ефимова, Антонова, 2012). Известно, что на освещенных участках число проростков может быть в два раза больше, чем на затененных (Дайнеко и др., 2017).
Сведения, приведенные в табл. 2, подтверждают влияние затенения. По мере развития заросли клена ясенелистного происходит смыкание крон, увеличение проективного покрытия и, как следствие, увеличение затенения. Из-за этого новые деревья начинают появляться преимущественно по периферии заросли, а не в ее глубине. Соответственно увеличивается степень сегрегации. Но рост этого показателя (С-критерия) не постоянен. В отдельные периоды может происходить уменьшение степени сегрегации старых и молодых деревьев. Это уменьшение может быть с двумя факторами – отмиранием наиболее старых деревьев внутри заросли клена ясенелистного и наличием позитивного влияния древесного полога на его прорастание. С отмиранием старых деревьев внутри заросли улучшаются условия освещения и увеличивается количество проростков. Позитивное влияние древесного полога на прорастание клена ясенелистного в плакорных условиях заключается в повышенном снегонакоплении внутри заросли, что повышает увлажнение почвы (Сапанов, 2009). До достижения определенной плотности деревьев положительное влияние повышенного снегонакопления превышает отрицательное влияние затенения на прорастание клена ясенелистного под древесным пологом. Для исследуемой территории при плотности до 750 деревьев на гектар молодые деревья появляются в основном под пологом заросли. При плотности свыше 750 деревьев на гектар они начинают появляться преимущественно за пределами заросли (см. рис. 3).
Заключение
В охранной зоне участка Ямская степь, у границы второго квартала, за 35 лет образовалась сомкнутая заросль клена ясенелистного. Процесс ее образования разделяется на три этапа. Первый этап (1980–1990 гг.) начался с появления первых деревьев и закончился образованием первого очага расселения клена ясенелистного вдоль межевой канавы. В это время деревья имели случайное пространственное распределение. Новые деревья появлялись преимущественно внутри первого очага расселения. Граница формирующейся заросли расширилась слабо, но росла плотность размещения деревьев внутри нее.
На втором этапе (1990–2000 гг.) заросль клена ясенелистного формируется уже вокруг двух отдельных очагов расселения, к которым в будущем будут приурочены участки максимальной плотности размещения деревьев. Деревья имеют групповое пространственное распределение, тенденция к группированию самая выраженная за весь анализируемый временной период. Новые деревья появляются преимущественно за пределами существующей границы заросли. В основном идет распространение клена ясенелистного вдоль межевой канавы, а также начинается расселение вбок от нее.
На третьем этапе (2000–2015 гг.) формируется сплошная сомкнутая заросль клена ясенелистного, охватывающая весь исследуемый участок. Деревья по-прежнему имеют групповое распределение в пространстве, но с каждым временным срезом оно все менее выраженное. Новые деревья появляются в основном в границах заросли.
Выявленные пространственные закономерности расселения клена ясенелистного вдоль линейных объектов (межевых канав) обусловлены сочетанием конфигурации территории, расположения источника заноса и расстояния массового ветрового переноса семян. Эти закономерности могут быть экстраполированы на территории, схожие с окрестностями Ямской степи по климатическим, геоморфологическим условиям и характеру землепользования. Такие условия наблюдаются на возвышенностях лесостепной зоны Восточно-Европейской равнины (Приднепровская, Среднерусская и Приволжская возвышенности).
Библиография
Агишев В. С. Таксономический состав пойменных прирусловых черноольшаников реки Боровка национального парка «Бузулукский бор» // Actualscience. 2017. Т. 3. № 1. С. 3–4.
Абрамова Л. М., Агишев В. С., Хазиахметов Р. М. Вселение клена ясенелистного (Acer negundo L., Asteraceae) в пойменные леса северо-запада Оренбургской области // Российский журнал биологических инвазий. 2019. Т. 12. № 2. С. 2–24.
Акатов В. В., Акатов Т. В., Гребенко Е. А. Изменения верхней границы распространения акации белой и клена ясенелистного в долине реки Белая (Западный Кавказ) // Лесоведение. 2014. № 1. С. 21–33.
Арбузова М. В. Древесные интродуценты в особо охраняемых природных территориях Белгородской области // Лесопользование, экология и охрана лесов: фундаментальные и прикладные аспекты: Материалы международной научно-практической конференции (г. Томск, 21–22 марта 2005 г.). Новосибирск: STT, 2005. С. 21–22.
Арбузова М. В., Украинский П. А. Клен ясенелистный (Acer negundo L.) в условиях Ямской степи (Белгородская область) // Флора и растительность Центрального Черноземья: Материалы межрегиональной научной конференции, посвященной Году особо охраняемых природных территорий и экологии (г. Курск, 8 апреля 2017 г.). Курск: Мечта, 2017. С. 134–135.
Арбузова М. В., Украинский П. А., Щербаков К. В. Древесно-кустарниковая растительность межевой канавы и прилегающей к ней территории Ямской степи // Флора и растительность Центрального Черноземья: Материалы межрегиональной научной конференции (г. Курск, 5 апреля 2014 г.). Курск: Мечта, 2014. С. 107–112.
Виноградова Ю. К., Майоров С. Р., Хорун Л. В. Черная книга флоры Средней России . М.: ГЕОС, 2010. 512 с.
Голеусов П. В., Артищев В. Е. Постселитебные геосистемы как ренатурационные элементы экологической сети Белгородской области // Современные проблемы ландшафтоведения и геоэкологии: Материалы VI Международной научной конференции (к 100-летию со дня рождения профессора В. А. Дементьева). Минск: БГУ, 2018. С. 116–119.
Дайнеко Н. М., Тимофеев С. Ф., Булохов А. Д., Панасенко Н. Н. Инвазия клена ясенелистного (Acer negundo L.) в условиях Добрушского района Гомельской области // Известия Гомельского государственного университета имени Ф. Скорины. 2017. № 3. С. 35–39.
Евстигнеев О. И., Мурашев И. А., Коротков В. Н. Анемохория и дальность рассеивания семян деревьев восточноевропейских лесов // Лесоведение. 2017. № 1. С. 45–52.
Емельянов А. В., Фролова С. В. Клен ясенелистный (Acer negundo L.) в прибрежных фитоценозах реки Ворона // Российский журнал биологических инвазий. 2011. Т. 4. № 2. С. 40–43.
Ефимова И. В., Антонова И. С. Развитие однолетних проростков Acer negundo L. в разных климатических и экологических условиях // Вестник Санкт-Петербургского университета. Сер. 3. Биология. 2012. № 3. С. 31–37.
Колтунова А. И., Кузьмин Н. И. Клен ясенелистный (Acer negundo L.) в Оренбуржье // Известия Оренбургского государственного аграрного университета. 2017. № 5. С. 211–213.
Костина М. В., Минькова Н. О., Ясинская О. И. Особенности клена ясенелистного в зеленых насаждениях Москвы // Российский журнал биологических инвазий. 2013. № 4. С. 32–43.
Леонтьев Д. Ф., Зверева К. А. Инвазия клена ясенелистного и облепихи по Московскому тракту на участке «Иркутск-Ангарск» // Бюллетень науки и практики. 2016. № 11. С. 40–44. DOI: 10.5281/zenodo.166777.
Рыжков О. В., Рыжкова Г. А. Использование ГИС-картографирования для изучения динамики растительного покрова пастбища Центрально-Черноземного заповедника и проектирования заповедно-режимных мероприятий // Режимы степных особо охраняемых природных территорий: Материалы международной научно-практической конференции, посвященной 130-летию со дня рождения профессора В. В. Алехина (г. Курск – пос. Заповедный, 15–18 января 2012 г.). Курск, 2012. С. 168–187.
Саксонов С. С. Инвазии Acer negundo L. (Aceraceae) в Ульяновской области // Самарская Лука: проблемы региональной и глобальной экологии. 2018. Т. 27. № 3. С. 215–219. DOI: 10.24411/2073-1035-2018-10072.
Сапанов М. К. Защитное лесоразведение в аридных регионах: исторический опыт и современные решения // Структура и функции лесов Европейской России. М.: Товарищество научных изданий КМК, 2009. С. 346–368.
Фатьянова Е. В., Лисицына О. В., Антонова И. С. Расселение клена ясенелистного (Acer negundo L.) в европейской лесостепи на примере некоторых географических пунктов Белгородской области и Республики Татарстан // Биоразнообразие: подходы к изучению и сохранению: Материалы Международной научной конференции, посвященной 100-летию кафедры ботаники Тверского государственного университета (г. Тверь, 8–11 ноября 2017 г.). Тверь: Тверской гос. ун-т, 2017. С. 418–420.
Abramson I. On bandwidth variation in kernel estimates – a square root law // Annals of Statistics. 1982. Vol. 10. № 4. P. 1217–1223.
Baddeley A., Rubak E., Turner R. Spatial point patterns: methodology and applications with R. London: Chapman and Hall/CRC Press, 2015. 810 p.
Bivand R. S., Pebesma E., Gómez-Rubio V. Applied Spatial Data Analysis with R. NY: Springer New York, 2013. 405 p. DOI: 10.1007/978-1-4614-7618-4.
Braun M., Schindler S., Essl F. Distribution and management of invasive alien plant species in protected areas in Central Europe // Journal for Nature Conservation. 2016. Vol. 33. P. 48–57. DOI: 10.1016/j.jnc.2016.07.002.
Camenen E., Porté A. J., Benito Garzón M. American trees shift their niches when invading Western Europe: evaluating invasion risks in a changing climate // Ecology and evolution. 2016. Vol. 20. № 6. P. 7263–7275. DOI: 10.1002/ece3.2376.
Clark P. J., Evans F. С. Distance to Nearest Neighbor as a Measure of Spatial Relationships in Populations // Ecology. 1954. Vol. 35. № 4. P. 445–453. DOI: 10.2307/1931034.
Dixon P. M. Nearest-neighbor contingency table analysis of spatial segregation for several species // Ecoscience. 2002. Vol. 9. № 2. P. 142–151. DOI: 10.1080/11956860.2002.11682700.
Dumitraşcu M., Bălteanu D. Invasive Terrestrial Plant Species in the Romanian Protected Areas: A Geographical Approach // Revue Roumaine de Géographie / Romanian Journal of Geography. 2014. Vol. 58. № 2. P. 145–160.
Marozas V., Cekstere G., Laivins M., Straigyte L. Comparison of neophyte communities of Robinia pseudoacacia L. and Acer negundo L. in the eastern Baltic Sea region cities of Riga and Kaunas // Urban Forestry & Urban Greening. 2015. Vol. 14. № 4. P. 826–834. DOI: 10.1016/j.ufug.2015.08.003.
Mędrzycki P. Inwazja amerykańskiego klonu Acer negundo L. a użytkowanie ziemi w Puszczy Białowieskiej / Praca doktorska. Warszawa: Uniwersytet Warszawski, 2002. 96 p.
R Core Team. 2020. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. URL: https://www.R-project.org.
Sachse U., Starfinger U., Kowarik I. Synanthropic woody species in the urban area of Berlin (West) // Urban ecology. The Hague: SPB Academic Publishing, 1990. P. 233–243.
Straigyte L., Cekstere G., Laivins M., Marozas V. The spread, intensity and invasiveness of the Acer negundo in Riga and Kaunas // Dendrobiology. 2015. Vol. 74. P. 157–168. DOI: 10.12657/denbio.074.016.
Valantinaite A., Straigyte L., Jurksiene G. Comparative analysis of invasion intensity of box elder (Acer negundo L.) and sosnowskyi hogweed (Heracleum sosnowskyj Manden) // Rural Development in Global Changes. 2011. Vol. 5. № 2. P. 161–166.
Благодарности
Исследование проведено при финансовой поддержке РФФИ (проект 18-35-20018 мол_а_вед).

 © 2011 - 2026
© 2011 - 2026