



Введение
Изучение пространственной структуры является одним из наиболее актуальных направлений в популяционной экологии позвоночных животных. Именно эта составляющая популяционной структуры служит, с одной стороны, основой всех форм нормальной жизнедеятельности вида, а с другой – самым лабильным и действенным механизмом приспособления к меняющимся условиям окружающей среды. Пространственная структура популяции объединяет широкий спектр особенностей использования территории и является показателем благополучия вида (Шилов, 1977).
Одним из наиболее удобных объектов исследований в этой области служат открыто живущие виды пустынных и полупустынных экосистем, в частности ящерицы. Их изучение становится все более насущной задачей в связи с зарастанием песков, которое встречается повсеместно на аридных территориях и влияет на популяции рептилий. В русле изучения динамики герпетокомплексов песчаных полупустынь Астраханской области (Полынова и др., 2019) одним из основных объектов наших исследований оказалась разноцветная ящурка Eremias arguta deserti (Gmelin, 1789) – фоновый вид соответствующих экосистем этого региона.
Численность разноцветной ящурки в разных частях ареала и различных биотопах колеблется в широком диапазоне от 1.2–3 до 550–640 ос/га (Разноцветная ящурка, 1993). Географически близкие материалы по Калмыкии 70-х – начала 80-х гг. прошлого века показывают относительную стабильность плотности населения ящурки в большинстве стаций (Бадмаева, 1983). В Астраханской области на данный момент происходит значительное сокращение численности вида.
Выяснение причин снижения численности разноцветной ящурки и изучение пространственной структуры как механизма приспособления к меняющимся условиям среды – цель нашего исследования. Объектом исследования является внутрипопуляционная группировка – уровень, на котором проходят начальные процессы авторегуляции в популяции.
Материалы
Материалы по пространственной структуре разноцветной ящурки собраны в течение трех полевых сезонов: в первые две декады мая 2017, 2018 гг. и в первой половине мая 2019 г. Вне сезона размножения – с 27 августа по 16 сентября 2017 г. и с 19 по 29 августа 2018 г. Место исследований – окрестности поселка Досанг Красноярского района Астраханской области (N 46° 54'08.7264" E 47° 54' 52.5312").
Выбор поселения вида стал результатом маршрутного обследования территории вокруг поселка (~7 км2). Выбранное поселение соответствует уровню внутрипопуляционной группировки или элементарной популяции (Наумов, 1963; Шилов, 1977) и расположено на отдельном участке полузакрепленного песка площадью 0.4 га. Фитоценоз – джузгуно-полынное сообщество (Calligonum aphyllum Litv. и Artemisia arenaria DC.) с наличием признаков сукцессии в сторону степного сообщества (Полынова и др., 2019).
Во время исследований на территории поселения отловлены, промерены и помечены все встреченные особи разноцветной ящурки: в мае и сентябре 2017 г. – 76 и 25 особей, в мае и конце августа 2018 г. – 38 и 14 особей, в мае 2019 г. – 12 ящериц.
Статья не содержит сведений по размерам индивидуальных участков, поскольку цель работы не требует обсуждения этого параметра пространственной структуры. Кроме того, представленные материалы в большинстве случаев характеризуют только центры активности участков обитания. У данного вида центры активности – это знакомые естественные убежища: участок с кустами и норами под ними. Центры активности легко выявляются за небольшой период наблюдений и дают представление о плотности поселения.
Методы
В работе использовался набор методов, подробно описанный нами в ранее опубликованных исследованиях (Полынова, Бажинова, 2012), поэтому здесь приведен лишь их краткий перечень.
Маркировку производили временной (спиртовой маркер) и постоянной (отрезание кончиков фаланг пальцев по классической схеме) меткой. У меченых животных измеряли длину тела и хвоста (мм). Описание пространственной структуры осуществляли на основе картирования встреч и перемещений, построения карт индивидуальных участков по методу выпуклого многоугольника, методов тропления и осторожного преследования. Обработка полевых данных проходила в программе Adobe llustrator (для визуализации участка). На основе обработанных материалов выявлена оседлая и мигрирующая части популяции. Характеристика численности дана в ос/га, а оценка величины потока мигрантов – в ос/сутки. Проведено геоботаническое исследование динамики фитоценоза (Полынова и др., 2019). Оценка зависимости экологических показателей дана на основе коэффициента корреляции Пирсона.
Результаты
Согласно опубликованным сведениям общий характер пространственного распределения популяций разноцветной ящурки неравномерен. На Прикаспийской низменности размещение мозаичное (Табачишин и др., 2006), а в Центральном Предкавказье и Харьковской области – ленточное (Разноцветная ящурка, 1993). В нашем случае поселение представляет элемент мозаичного распределения, типичного для региона.
Трехлетние наблюдения, проведенные на уровне внутрипопуляционной группировки, свидетельствуют о направленном снижении численности вида. Значительно уменьшается как оседлая часть поселения, так и поток мигрантов (табл. 1).
Таблица 1. Динамика плотности населения группировки разноцветной ящурки Eremias arguta deserti,
май 2017–2019 гг.
| Сезон | Общая плотность, ос/га | Плотность оседлых особей, ос/га | Поток мигрантов, ос/сутки |
| Май 2017 | 190 | 85 | 2.1 |
| Май 2018 | 95 | 17.5 | 1.5 |
| Май 2019 | 30 | 7.5 | 0.5 |
Процесс затрагивает не только количественную, но и качественную характеристику группировки: происходят принципиальные перемены в пространственной структуре и половозрастном составе.
Разноцветная ящурка, как большинство ящериц (Stamps, 1977; Vitt et al., 1974 и др.), относится к животным с интенсивным типом использования территории. Животные образуют систему групп перекрывающихся участков, размер которых колеблется в широком диапазоне (Тертышников, 1970). Главной функцией подобной пространственной структуры в сезон размножения является поддержание достаточного уровня контактов между брачными партнерами. Такую пространственную структуру группировки мы наблюдаем у разноцветной ящурки весной 2017 г. Это система перекрывающихся участков брачных партнеров (рис. 1) при соотношении половых групп 1 : 1.1 (16 самцов : 18 самок).
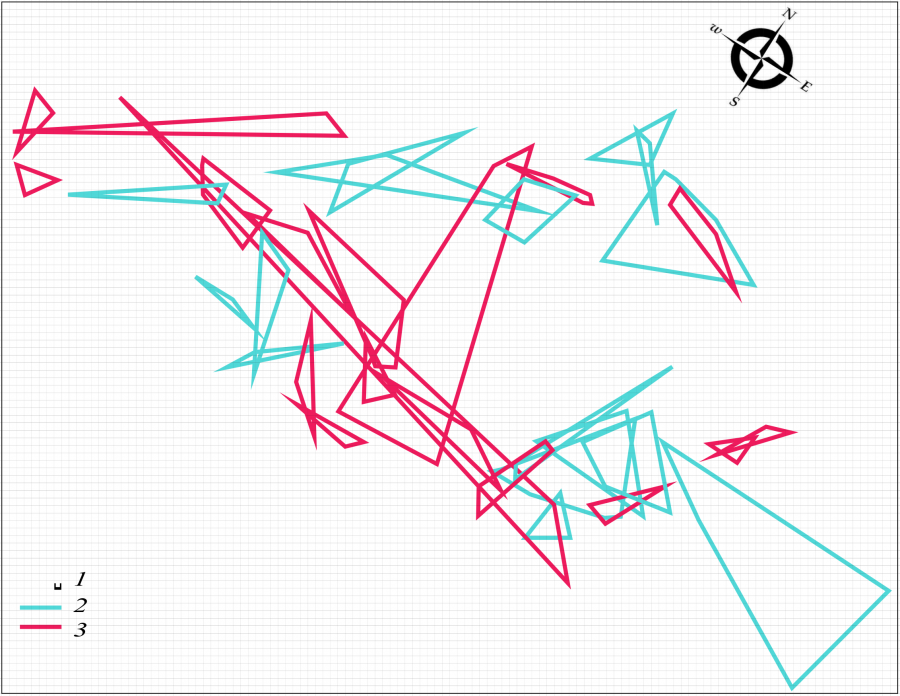
Рис. 1. Пространственная структура внутрипопуляционной группировки разноцветной ящурки Eremias arguta deserti, май 2017 г. 1 – размер ячейки 1 м, 2 – граница участка самца, 3 – граница участка самки
Fig. 1. Spatial structure of intra-population group of the stepperunner Eremias arguta deserti, May 2017. 1 – size of cell is 1 m, 2 – boundary of the male area, 3 – boundary of the female area
Система участков самок объединяет между собой всю внутрипопуляционную группировку. Неполовозрелых особей в группировке нет. Поток мигрантов, проходящий по территории поселения (см. табл. 1), также состоит только из половозрелых ящериц (18 самцов, 24 самки). Под потоком мигрантов мы понимаем не направленно движущихся животных, а особей, широко кочующих по территории популяции и не имеющих в данном месте постоянного участка.
Весной 2018 г. пространственная структура поселения разноцветной ящурки существенно меняется. Оседлая часть группировки также состоит из самцов и самок (2 и 5 соответственно), но уменьшается в 4.8 раза (см. табл. 1). Кроме того, участки половых партнеров располагаются изолированно (рис. 2). Подобная пространственная структура гораздо меньше способствует контакту брачных партнеров. Как и весной предыдущего года, неполовозрелых ящериц среди оседлых особей нет. В небольшом числе они встречаются среди кочующих животных, составляя 10 % от общего числа мигрантов (11 самцов, 17 самок, 3 неполовозрелые особи). Поток мигрантов также снижается (см. табл. 1), что подтверждает снижение численности популяции в целом.
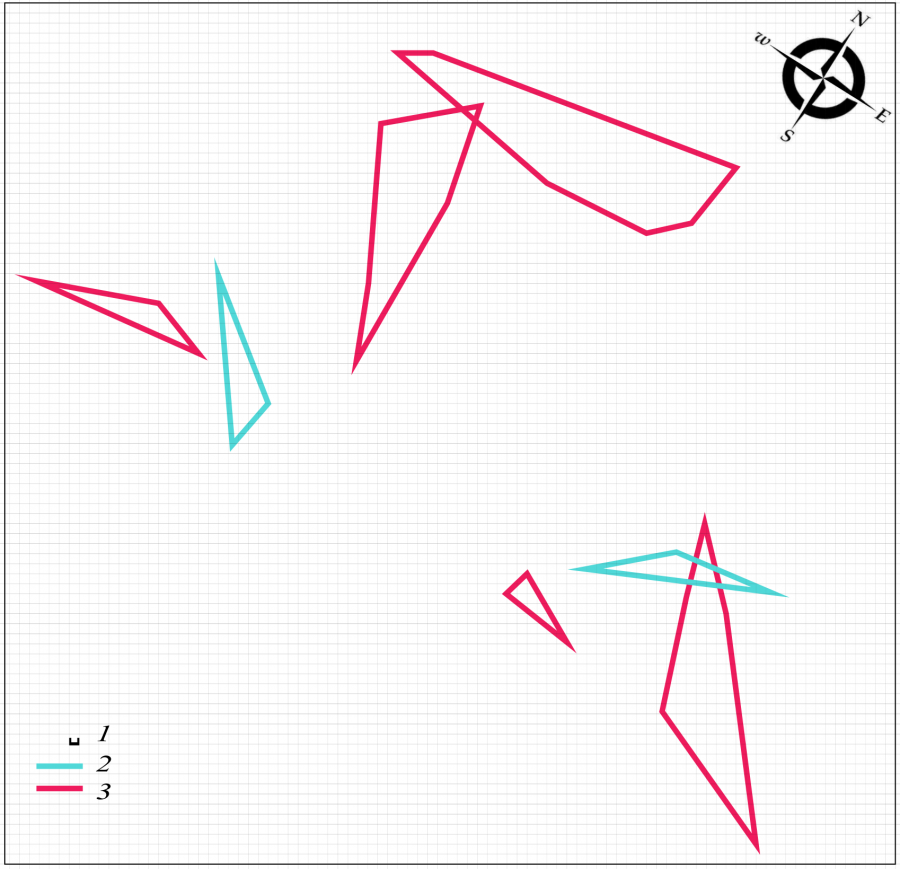
Рис. 2. Пространственная структура внутрипопуляционной группировки разноцветной ящурки Eremias arguta deserti, май 2018 г. 1 – размер ячейки 1 м, 2 – граница участка самца, 3 – граница участка самки
Fig. 2. Spatial structure of intra-population group of the stepperunner Eremias arguta deserti, May 2018. 1 – size of cell is 1 m, 2 – boundary of the male area, 3 – boundary of the female area
Весной 2019 г. от группировки разноцветной ящурки на данной территории остается 3 изолированных участка самок (рис. 3). Оседлых самцов нет. При таком составе функцию поддержания достаточного уровня брачных контактов пространственная структура, очевидно, уже не выполняет. Животных значительно меньше и в кочующей части поселения (табл. 1), что является показателем продолжающегося снижения общей численности популяции. Неполовозрелые ящерицы на территории поселения по-прежнему отсутствуют.
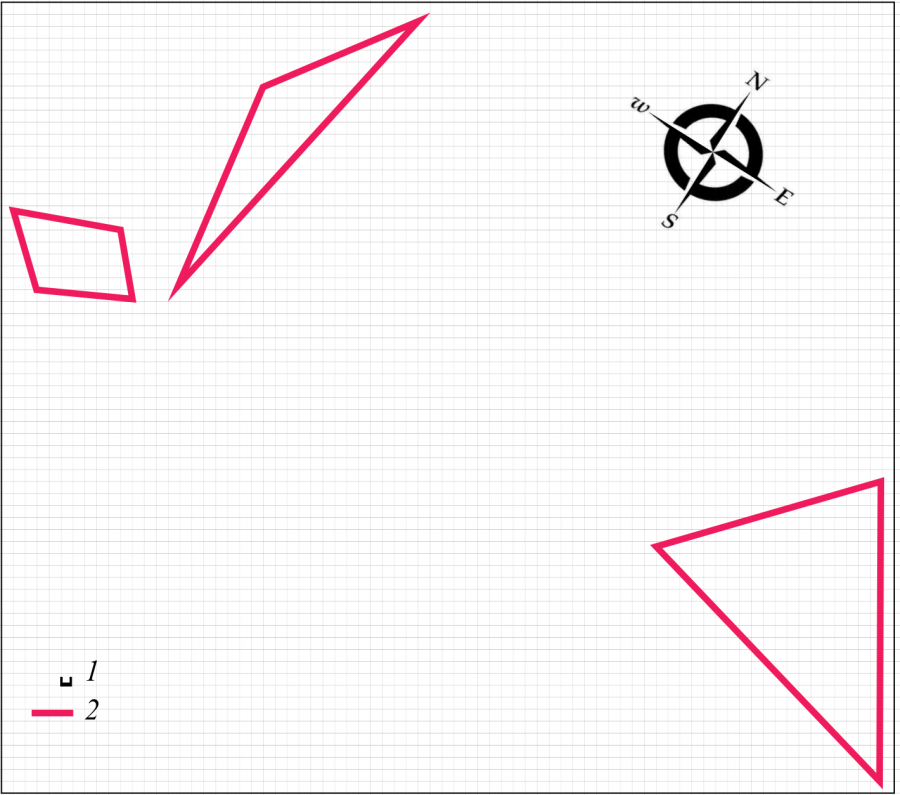
Рис. 3. Пространственная структура внутрипопуляционной группировки разноцветной ящурки Eremias arguta deserti, май 2019 г. 1 – размер ячейки 1 м, 2 – граница участка самки
Fig. 3. Spatial structure of intra-population group of stepperunner Eremias arguta deserti, May 2019. 1 – size of cell is 1 m, 2 – boundaryof the female area
Показателем успешности брачного сезона сокращающейся группировки служит выход сеголеток в конце лета – начале осени. В 2017 г. на территории группировки мы встречаем 18 сеголеток, а в 2018 г. – 10, т. е. в пересчете на плотность населения – 45 и 25 ос/га соответственно. Снижение численности и в этой возрастной группе очевидно.
Основной причиной уменьшения численности вида, на наш взгляд, являются изменения характерного биотопа. По нашим наблюдениям, которые проходили в полевые сезоны 2010–2014 и 2017–2019 гг., происходит зарастание местообитания внутрипопуляционной группировки и примыкающей к ней территории популяции. В упрощенном варианте геоботанического исследования использованы два параметра: видовое разнообразие фитоценоза и общее проективное покрытие травяного яруса. Проведенное в 2011, 2014 и 2017 гг. геоботаническое описание площадок достоверно показывает, что за рассмотренный период слабо закрепленная территория превращается в полузакрепленную, полузакрепленная – в закрепленную, а общее проективное покрытие травяного яруса закрепленного участка увеличивается в 1.6 раза (Полынова и др., 2019). В 2018 и 2019 гг. во флоре к 13 постоянно встречающимся видам прибавляется еще 10 новых однолетников и двулетников (Мишустин, Полынова, 2019а, б). С 2011 г. мы также наблюдаем за распространением колосняка гигантского Leymus racemosus (Lam.) Tzvelev – основного участника закрепления песчаных экосистем (Цвелев, 1976). При геоботаническом описании территории весной 2011 г. растение фиксируется на 1/4 всех заложенных площадок, весной 2014 – на 1/3, а весной 2017 г. колосняк присутствует уже на половине площадок. Сукцессионный процесс приводит к изменению джузгуно-полынного сообщества на джузгуно-разнотравное, т. е. идет в сторону остепнения биоценоза (Полынова и др., 2019).
Анализ динамики основных климатических факторов исследуемой территории показывает, что зарастание песков связано с увеличением общей суммы осадков (Архив погоды в Досанге, 2020). Результат расчета линейного коэффициента корреляции Пирсона свидетельствует (r = 0.82; p ≤ 0.01), что в течение последнего десятилетия увеличение общего проективного покрытия растительного яруса имеет сильную положительную зависимость от увеличения общей суммы осадков (рис. 4).
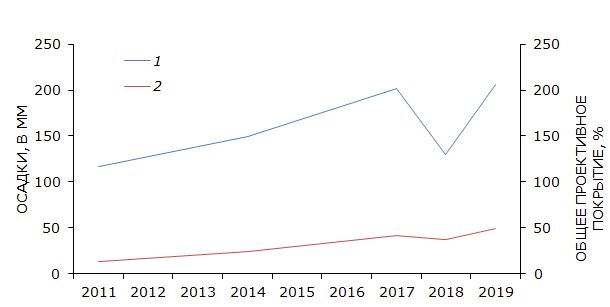
Рис. 4. Многолетняя динамика общей суммы осадков и проективного покрытия. 1 – общая сумма осадков (мм), 2 – общее проективное покрытие травяного яруса (%)
Fig. 4. Long-term dynamics of the total amount of precipitation and total projective cover of vegetation layer. 1 – total amount of precipitation (mm); 2 – total projective cover of vegetation layer (%)
Обсуждение
Снижение численности многих видов рептилий является широко распространенным явлением на зарастающих территориях песчаных пустынь и полупустынь нашей страны и сопредельных государств: Поволжья (Табачишин и др., 2006), Казахстана (Сараев, Пестов, 2010), Восточного Предкавказья (Лотиев, Батхиев, 2019), Узбекистана (Бондаренко и др., 2010). Подобный процесс отмечен и в условиях зарастания биотопов с каменистым грунтом (Galan, 2004). Процесс зарастания зависит прежде всего от климатических условий (Колесниченко, 1971). Наши материалы показывают его положительную корреляцию с увеличением общей суммы осадков за последнее десятилетие. Закреплению песков также способствует повсеместное сокращение поголовья скота и исчезновение диких копытных, отмеченное и для исследованной территории.
Изменение характерного биотопа негативно влияет прежде всего на виды псаммофилы (Полынова и др., 2019), также оказывая отрицательное воздействие на экологически пластичную разноцветную ящурку. Разноцветная ящурка относится к эврибионтным видам, однако имеет определенные стации предпочтения в разных зонах, что приводит к сложной картине биотопического распределения (Разноцветная ящурка, 1993). Выбор биотопа у разноцветной ящурки зависит от целого ряда факторов. Важную роль играет освещенность, что определяется структурой растительного покрова. На песчаных почвах предпочитает горизонтальные поверхности и участки с относительно разреженным травостоем (Котенко, 1983). Высокая численность ящерицы установлена в биотопах с проективным покрытием от нескольких до 30–40 % (Разноцветная ящурка, 1993). По нашим наблюдениям, негативными последствиями зарастания биотопа для вида являются ухудшения условий передвижения и коммуникации. Наиболее предпочитаемые стации вида имеют низкую влажность, песчаный или каменисто-песчаный грунт, высокую освещенность, возможность передвигаться и достаточное количество убежищ (Разноцветная ящурка, 1993).
На территории исследований вид селится на относительно открытых участках, образуя на них систему внутрипопуляционных группировок, пространственная и половозрастная структура которых реагирует на зарастание биотопа. В начале исследований, весной 2017 г., мы наблюдаем на такой открытой территории многочисленную группировку. Значительное перекрывание участков самцов и самок и их соотношение, близкое к 1 : 1, создают основу для процесса размножения. Подобное соотношение полов характерно для популяций разноцветной ящурки из разных частей ареала (Разноцветная ящурка, 1993) и свойственно многим видам ящериц в сезон размножения (Alcala, Brown, 1967; Bustard, 1970; Ballinger, 1973 и др.).
В следующий брачный сезон 2018 г. группировка продолжает свое существование на той же территории, адаптируясь к изменившимся условиям. Число оседлых животных резко уменьшается. Стремясь занять пространство с разреженной растительностью, животные образуют структуру с почти не перекрывающимися индивидуальными участками. Контакты брачных партнеров в этой ситуации становятся более редкими, что снижает эффективность процесса размножения. В результате численность сеголеток в конце августа уменьшается по сравнению с предыдущим годом в 1.8 раза. Снижение потока мигрантов свидетельствует о падении численности популяции в целом.
В весенний сезон 2019 г. способной к размножению группировки на территории поселения нет. Оседлы только немногочисленные изолированно живущие самки, занявшие участки с разреженной растительностью. Неоседлая часть поселения продолжает сокращаться. Популяция постепенно перестает размножаться на данной территории, что подтверждается отсутствием неполовозрелых особей во все весенние сезоны и снижением численности сеголеток в 2018 г. по сравнению с 2017 г.
Таким образом, на зарастание характерного биотопа группировка отвечает поэтапным снижением численности за счет постепенного сокращения размножения в измененных условиях. Снижение численности вида на данной территории подтверждено и наблюдениями наших коллег (Литвинов Н. А., устное сообщение).
Заключение
- Трехлетнее исследование пространственной структуры разноцветной ящурки Eremias arguta deserti (Gmelin, 1789) показывает направленное сокращение численности популяции, которое наглядно проявляется на уровне внутрипопуляционной группировки.
- Пространственная структура группировки в брачный сезон упрощается от сезона к сезону. Из системы перекрывающихся участков самцов и самок она сначала трансформируется в систему изолированных участков половых партнеров, а затем в малочисленную группировку изолированных участков самок. Это свидетельствует о том, что размножение на данной территории постепенно сокращается. Уменьшение мигрирующей части поселения подтверждает общее сокращение численности популяции.
- Основной причиной описанного процесса является изменение характерного биоценоза: происходит увеличение общего проективного покрытия травяного яруса, разрастание закрепляющих пески видов растений и общее остепнение растительных сообществ. Протекающие в экосистеме изменения связаны с увеличением общей суммы осадков на данной территории за последнее десятилетие. Негативными последствиями зарастания биотопа для вида являются ухудшения условий передвижения и коммуникации.
Библиография
Архив погоды в Досанге . URL: https://rp5.ru/Архив_погоды_в_Досанге (дата обращения: 23.01.2020).
Бадмаева В. И. Ящерицы Калмыкии : Дис. … канд. биол. наук. Киев, 1983. 227 с.
Бондаренко Д. А., Перегонцев Е. А., Абдуназаров Б. Б., Сударев В. О. О современном состоянии герпетофауны песчаных массивов Ферганской долины (Узбекистан) // Герпетологические исследования в Казахстане и сопредельных странах: Сборник научных статей / Под ред. Т. Н. Дуйсебаевой. Алматы: АСБК СОПК, 2010. С. 246–248.
Котенко Т. И. Пресмыкающиеся левобережной степи Украины : Дис. … канд. биол. наук. Киев, 1983. 555 с.
Лотиев К. Ю., Батхиев А. М. О деградации туранского герпетофаунистического комплекса в Терском песчаном массиве (Восточное Предкавказье) // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2. С. 115–128. DOI: 10.21685/2307-9150-2019-2-12.
Мишустин С. С., Полынова Г. В. Влияние абиотических факторов на популяции ящериц в условиях Астраханских полупустынь // Современная герпетология: проблемы и пути их решения. СПб.: Зоологический институт РАН, 2019а. С. 61.
Мишустин С. С., Полынова Г. В. Динамика фитоценозов в полупустынях юго-восточной части Нижнего Поволжья // Вестник института ИКИАТ. 2019б. № 1 (38). С. 10–11. DOI: 10.24411/2071-7830-2019-10002.
Наумов Н. П. Экология животных . М.: Высшая школа, 1963. 618 с.
Полынова Г. В., Бажинова А. В. Учет особенностей активности при оценке численности популяции круглоголовки-вертихвостки (Phrynocephalus guttatus guttatus) // Зоологический журнал. 2012. Т. 91. № 11. С. 1411–1414.
Полынова Г. В., Мишустин С. С., Полынова О. Е. Динамика герпетокомплекса песчаных пустынь Астраханской области // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2. С. 150–163. DOI: 10.21685/2307-9150-2019-2-15.
Разноцветная ящурка : Коллективная монография / Под ред. Н. Н. Щербака. Киев: Наукова думка, 1993. 240 с.
Сараев Ф. А., Пестов М. В. К кадастру рептилий Северного и Северо-Восточного Прикаспия // Герпетологические исследования в Казахстане и сопредельных странах: Сборник научных статей / Под ред. Т. Н. Дуйсебаевой. Алматы: АСБК – СОПК, 2010. С. 172–191.
Табачишин В. Г., Завьялов Е. В., Табачишина Е. И. Пространственное размещение разноцветной ящурки – Eremias arguta (Pallas, 1776) на севере ареала в Поволжье // Современная герпетология. 2006. Т. 5/6. С. 117–124.
Тертышников М. Ф. Индивидуальная территория прыткой ящерицы (Lacerta agilis exigua Eichw., 1831) и разноцветной ящурки (Eremias arguta deserti Gmel., 1876) и особенности ее использования // Зоологический журнал. 1970. Т. 49. Вып. 9. С. 1377–1385.
Цвелев Н. Н. Злаки СССР . Л.: Наука, 1976. 788 с.
Шилов И. А. Эколого-физиологические основы популяционных отношений у животных . М.: Московский университет, 1977. 261 с.
Alcala A. C., Brown W. C. Population ecology of the tropical scincoid lizard, Emoia atrocostata in the Philippines // Copeia. 1967. No 3. P. 596–604.
Ballinger R. E. Comparative demography of two viviparous iguanid lizards (Sceloporus jarrovi and Sceloporus poinsetti) // Ecology. 1973. Vol. 54. No 2. P. 269–283.
Bustard H. R. The population ecology of the Australian gekkonid lizard Heteronica binoei in an exploited forest // Journal of zoology. 1970. Vol. 162. No 1. P. 31–42.
Galan P. Structure of a population of the lizard Podarcis bocagei in northwest Spain: variations in age distribution, size distribution and sex ratio // Animal biology. 2004. Vol. 54. No 1. P. 57–75.
Stamps J. F. Social behavior and special pattern in lizards // Biology of the reptilian. New York: Akad. Press, 1977. Vol. 7. P. 265–334.
Vitt L. J., Congdon J. D., Hulse A. C., Platz J. K. Territorial aggression encounters and tail breaks in the lizard Sceloporus magister // Copeia. 1974. No 4. P. 990–993.
Благодарности
Публикация подготовлена при поддержке «Программы РУДН 5-100».

 © 2011 - 2026
© 2011 - 2026