



Введение
Будучи немногочисленным, но достаточно широко распространенным видом мелких млекопитающих Палеарктики, равнозубая бурозубка по-прежнему остается одним из наименее изученных представителей местной фауны. Объясняется это не только ее малочисленностью, но и скрытным образом жизни, что вызывает известные трудности ее изучения. К тому же в качестве самостоятельного вида (и, соответственно, отдельного объекта исследований) равнозубая бурозубка была выделена из номинального таксона сравнительно недавно – лишь в конце 1960-х гг. (Долгов, 1985). В итоге, если для большинства других видов бурозубок не составляет особого труда назвать по крайней мере десяток специальных монографий и чуть ли не сотню посвященных им специальных обзорных статей, то для равнозубой все ограничивается максимум несколькими специальными публикациями, при этом относящимися в основном к территории Северо-Востока Сибири (Докучаев, 1981, 1990).
В этой связи настоящее сообщение имеет целью восполнить образовавшийся пробел в отношении европейской области распространения равнозубой бурозубки путем обобщения всех накопленных к настоящему времени собственных и литературных данных по биологии этого вида в условиях северо-западной периферии ареала.
Материалы
Материалом для настоящей работы послужили многолетние сборы зверьков рассматриваемого вида, отлавливаемых двумя основными методами: ловушко-линиями (плашки Геро) и ловчими 30-метровыми канавками. Первым было отработано 360270 ловушко-суток и добыто 90 равнозубых бурозубки, вторым – 9180 канавко-суток и 97 зверьков. В итоге общее число добытых и проведенных через зооанатомический анализ особей рассматриваемого вида составило 187. Что же касается проведенных исследований – оценки численности и территориального размещения зверьков, анализа репродуктивной системы и хода репродукции, а также определения возрастного и полового состава популяции, изучения линьки шерстного покрова и особенностей питания, то все это проводилось по общепринятой методике (обзоры см.: Новиков, 1953; Карасева и др., 2008; Ивантер, 1975, 2018).
Результаты
Численность и биотопическое размещение
Равнозубая бурозубка – типичный восточный палеаркт с ареалом, простирающимся от Финляндии до Камчатки. Обычна она на Дальнем Востоке и в Сибири (Долгов, 1968; Юдин, 1971), но в европейской части России численность ее, как правило, низка. Не составляет исключения в этом отношении и Карелия, где данный вид относится к числу редких и занимает по численности одно из самых последних мест. Его доля в уловах давилками равна 0.5 % от общего числа Micromammalia и 5.4 % от добытых Soricidae, канавками — 0.7 % и 0.9 % соответственно. Средняя численность 0.024 экз. на 100 ловушко-суток и 0.1 на 10 канавко-суток. На весьма ограниченном материале невозможно проследить движение численности по годам, однако то, что большую часть его составляют зверьки, добытые в 1968—1970 гг. (25 экз., т. е. почти 70 %), позволяет предположить наличие в этот период значительного подъема численности равнозубой бурозубки.
Ареал этого вида включает, по-видимому, всю Восточную Фенноскандию, но, как уже указывалось, на этой территории она малочисленна или редка. По характеру биотопического размещения сибирская форма равнозубой бурозубки наиболее тесно связана с таежными формациями. В условиях же Северо-Запада России она заселяет разнообразные биотопы, но наиболее часто добывается в ландшафтах с наличием крупных массивов спелых (особенно еловых и хвойно-лиственных) лесов, в зрелых лиственных древостоях с хорошо выраженным травяным покровом, а также на захламленных и поросших молодым лесом участках луговин. Относительная стенотопность и тяготение равнозубой бурозубки к закрытым лесным биотопам таежного типа – темнохвойным лесам и их производным (хвойно-лиственным насаждениям и зарастающим гарям), долинным ельникам и в меньшей степени вторичным березово-осиновым лесам), по-видимому, вообще характерная черта, поскольку и в других частях ареала она проявляется достаточно четко (Юдин, Барсова, 1967; Долгов и др., 1968; Ревин, 1968; Пучковский, 1969, 1973, 2000; Попов, 1971; Лукьянова, 1975, 1992; Равкин, Лукьянова, 1976; Куприянова, 1978, 1994; Юдин и др., 1979; Докучаев, 1990, 1994; Большаков и др., 1996; Бобрецов др., 2004; Skaren, 1964; Siivоnen, 1965). Открытых пространств с задернованной плотной почвой, сухих чистых сосняков и болот равнозубая бурозубка избегает. То же относится и к лесосекам, так что среди всех представителей Soricidae равнозубая бурозубка демонстрирует самую негативную реакцию на вырубку хвойных лесов.
Территориальная структура популяции и индивидуальные участки
В плане территориальности средняя бурозубка изучена совершенно недостаточно. По данным Н. В. Моралевой (1983, 1987), в пойме Енисея (ивово-ольховые лесные и кустарниковые ассоциации) этот вид по сравнению с другими представителями рода Sorex продемонстрировал на площадке мечения самую низкую степень оседлости (повторно было поймано только около 10 % меченых зверьков). Естественно, что столь незначительная доля возвратов не позволила сколько-нибудь полно оценить особенности индивидуального использования территории, характерные для данного вида. В связи с этим представляет интерес и тот сравнительно небольшой материал, который был получен при индивидуальном мечении равнозубых бурозубок в карельском Приладожье (Макаров, 1990; Ивантер, Макаров, 2001). Численность этого вида в районе исследований была весь период наблюдений низка, и в целом в уловах землероек на него приходилось всего около 1.1 %.
За весь период живоотлова было помечено 11 особей равнозубых бурозубок (из них 10 — в 1989 г.). Общее количество поимок – 77. Повторно отлавливались 10 зверьков (91 %). Максимальное число поимок одной особи – 17. В дальнейшем основной анализ идет только пикового 1989 г.
Все меченые животные концентрировались на территории около 1 га в пределах одной экспериментальной площадки, отдавая явное предпочтение перестойному ельнику с большим количеством бурелома и богатым напочвенным покровом из папоротников с кислицей и малиной. На данный биотоп, занимающий около 18 % территории площадок, пришлось 84 % поимок равнозубых бурозубок. Подобная биотопическая приуроченность согласуется с данными, полученными отловом канавками и давилками (Ивантер, Макаров, 2001). Тесная связь данного вида с лесными стациями, а прежде всего с темнохвойными лесами, отмечается на большей части ареала (Долгов, 1985). Отчетливое тяготение равнозубой бурозубки к крупнотравным приручьевым и зеленомошным ельникам выявлено в средней тайге Коми (Куприянова, Ануфриев, 1992) и в Карелии (Ивантер, 1975). Связь данного вида как наиболее крупного здесь по размерам представителя рода Sorex с наиболее продуктивными биоценозами объясняется высокими показателями биомассы обитающих здесь напочвенных и почвенных беспозвоночных (Hanski, Kaikusalo, 1989).
Агрегированность особей данного вида на территории одного биотопа, отмеченная на площадке мечения, может одновременно служить и средством поддержания определенной плотности, необходимой для размножения вида с низкой численностью. По заселенной площади зверьки распределяются сравнительно равномерно; из посещавшихся зверьками живоловок лишь в 2 ловилось по три особи, в 12 – по две и в 32 – по одной.
Для оценки взаимосвязи распределения особей доминирующей по численности обыкновенной бурозубки и редкой равнозубой бурозубки был рассчитан коэффициент корреляции поимок особей этих видов (Ивантер, 2018). Предварительно можно было предположить, что коэффициент, близкий к –1, будет означать либо жесткие конкурентные взаимоотношения в какой-либо форме, либо существенные различия в экологических требованиях видов, а к +1, напротив, сходство экологических потребностей. В результате же реального расчета был получен коэффициент 0.05, который означает, что распределение двух видов независимо друг от друга. Таким образом, предположение о возможном вытеснении рассматриваемого вида из субоптимальных биотопов доминирующей обыкновенной бурозубкой (Лукьянова, 1992) на нашем материале не подтвердилось.
Все зверьки были помечены в начальный цикл отловов в июне (66 поимок), в августе на площадке наблюдалось только 3 особи, новых не появилось. Возрастно-половой состав популяции за время наблюдений несколько изменился: в июле на площадке было 6 зимовавших зверьков и 4 сеголетка, в августе отмечен 1 зимовавший самец и 2 сеголетка. Высокая доля зимовавших особей в популяции отмечалась и ранее по результатам канавочных и давилочных отловов, что свидетельствует о низкой интенсивности размножения вида (Ивантер, Ивантер, 1988).
Площади индивидуальных участков зимовавших самцов составили в июле 1200, 1700, 2300 и 2600 м2 (последняя особь отмечалась и в августе, когда площадь индивидуального участка составила 1900 м2). Среднее количество попадаемости одной особи 10.7. При этом частота поимок в центре участков наибольшая, а расстояние между крайними точками отловов достигает 130–140 м.
Точно оценить размер индивидуальных участков зимовавших самок не удалось: одна особь обитала на границе площадки мечения, вторая повторно не отлавливалась. Измеренные участки обитания сеголетков составили 700, 1000 и 1400 м2, максимальное расстояние между точками отловов – 100 м.
За месяц границы индивидуальных участков зверьков значительно изменились, но все они либо сохранили общие территории со своими июльскими участками, либо граничили с ними (рис. 1). Изменение участков пошло в направлении площадей, где в августе другие особи этого вида не отлавливались, но это происходило в пределах биотопов, занятых популяцией в июле.
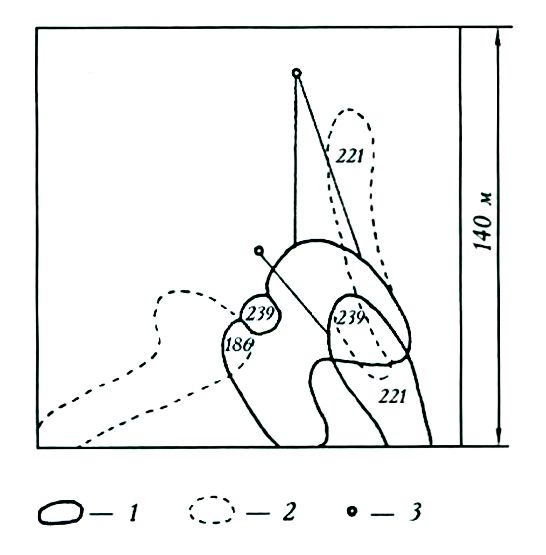
Рис. 1. Индивидуальные участки равнозубых бурозубок в июле (1) и августе (2), 3 – отдельные поимки зверьков (№ 186 и 221) в июле вне индивидуальных участков
Fig. 1. Home ranges of equal-toothed shrews in July (1) and in August (2), 3 – separate catches (№ 186 and 221) in July outside home ranges
Таким образом, в Карелии, по крайней мере в условиях коренных темнохвойных лесов, равнозубые бурозубки ведут оседлый образ жизни. Зверьки эти подвижнее, чем обитающие здесь же обыкновенные бурозубки, но индивидуальные участки у них более стабильны во времени и пространстве. Размеры участков сеголеток больше, чем у обыкновенной бурозубки. Большую оседлость, в сравнении с аналогичной группой обыкновенных бурозубок, демонстрируют и зимовавшие самцы.
Особенности питания
Питание равнозубой бурозубки изучено слабо, что связано с невысокой ее численностью на большей части ареала (Долгов, 1985), а также трудностями технического характера. Особенности питания этого вида в природе изучались в Западном Предверхоянье (Вольперт, Аверенский, 1983; Вольперт, 1986), в Карелии (Ивантер и др., 1973; Ивантер, Ивантер, 1988; Ивантер, Макаров, 2001), в Зейском заповеднике (Бромлей и др., 1984) и на юге Приморья (Нестеренко, 1999). Объем исследованного материала во всех случаях был невелик, что осложняет понимание географических различий трофических связей этого вида. Питание равнозубой бурозубки изучалось также при содержании в неволе (Охотина, 1974).
Как показали предшествующие исследования, основу рационов зверьков составляют жесткокрылые (их содержат 50–70 % желудков) и дождевые черви (55–68 %) (Ивантер и др., 1973; Вольперт, Аверенский, 1983). Существенной в питании может быть доля личинок чешуекрылых (до 40 %) (Ивантер и др., 1973), двукрылых (до 44 %) (Вольперт, Аверенский, 1983), паукообразных (до 50 %) (Ивантер, Ивантер, 1988). В неволе охотно едят имаго и личинок прямокрылых, дождевых червей, перепончатокрылых, муравьиных куколок и личинок жесткокрылых (Охотина, 1974), при этом доля дождевых червей в рационе уменьшается, что, вероятно, связано с наличием других, более предпочитаемых, кормов. Растительный материал в питании значительной роли не играл (до 11 %), однако в неволе отмечалось поедание кедровых орешков (Лукьянова, 1974).
Эффективность использования пищи равнозубой бурозубкой меньше, чем у других североевропейских Sorex, за исключением крошечной бурозубки (Hanski, 1984).
Находящийся в нашем распоряжении материал по питанию равнозубой бурозубки получен в результате разбора содержимого 10 желудков зверьков из заповедника «Кивач» (табл. 1) и 43 желудков из Приладожья (табл. 2). Согласно этим данным, основу рациона рассматриваемого вида составили насекомые (88 %), дождевые черви (63 %) и паукообразные (49 %). Из насекомых чаще других поедались личинки двукрылых (37 %) и чешуекрылых (28 %), а также жесткокрылые (47 %). Последние были представлены жужелицами и стафилинами. Из паукообразных в питании преобладали сенокосцы, доля пауков вдвое меньше. Полученные данные вполне согласуются с более ранними результатами, отличия состоят в основном в меньшем потреблении жесткокрылых и большем потреблении паукообразных.
Таким образом, рацион равнозубой бурозубки носит явно выраженный почвенный характер – в нем преобладают почвенные и напочвенные организмы, единственное исключение – гусеницы чешуекрылых.
Таблица 1. Состав пищи равнозубой бурозубки (по 10 желудкам, Кивач, июль – октябрь 1959 г.)
Table 1. Food composition of the equal-toothed shrew (by 10 stomaches, “Kivach”, July – October 1959)
| Вид корма | Абс. | Число встреч, % от числа исследованных желудков |
| Насекомые | 8 | 80.0 |
| Насекомые, ближе не определенные | 2 | 20.0 |
| Личинки насекомых, ближе не определенные | 3 | 30.0 |
| Яйца насекомых | 1 | 10.0 |
| Жесткокрылые | 7 | 70.0 |
| в том числе: | ||
| имаго | 7 | 70.0 |
| личинки | 2 | 20.0 |
| Щелкуны (личинки) | 1 | 10.0 |
| Жужелицы (имаго) | 1 | 10.0 |
| Чешуекрылый (гусеницы) | 4 | 40.0 |
| Паукообразные | 1 | 10.0 |
| Многоножки | 1 | 10.0 |
| Дождевые черви | 6 | 60.0 |
| Растительные остатки | 1 | 10.0 |
Таблица 2. Состав пищи равнозубой бурозубки в Приладожье (встречаемость, % к общему числу желудков с идентифицированными объектами питания)
Table 2. The food composition of the equal-toothed shrew in Ladoga region (occurence, % of the total number of stomachs with identified food items)
| Вид корма | Абс. | Встречаемость, % |
| 1 | 2 | 3 |
| Насекомые (Insecta) | 38 | 88.4 |
| имаго | 17 | 39.5 |
| личинки | 25 | 58.1 |
| куколки | 3 | 7.0 |
| яйца | 1 | 2.3 |
| 1 | 2 | 3 |
| Жесткокрылые (Coleoptera) | 20 | 46.5 |
| имаго | 16 | 37.2 |
| личинки | 10 | 23.3 |
| Carabidae | 6 | 14.0 |
| имаго | 5 | 11.6 |
| личинки | 1 | 2.3 |
| Staphylionidae | 5 | 11.6 |
| имаго | 3 | 7.0 |
| личинки | 3 | 7.0 |
| Elaleridae (личинки) | 3 | 7.0 |
| Curculionidae (имаго) | 2 | 4.7 |
| Жесткокрылые, ближе не определенные | 12 | 27.9 |
| имаго | 7 | 16.3 |
| личинки | 5 | 11.6 |
| Двукрылые (Diptera) | 18 | 41.9 |
| имаго | 5 | 11.6 |
| личинки | 16 | 37.2 |
| Bibionidae (личинки) | 2 | 4.7 |
| Sciaridae (личинки) | 6 | 14.0 |
| Tipulidae (личинки) | 3 | 7.0 |
| Rhagiomdac (личинки) | 1 | 2.3 |
| Mycelophilidae (личинки) | 1 | 2.3 |
| Двукрылые, ближе не определенные | 11 | 25.6 |
| имаго | 5 | 11.6 |
| личинки | 8 | 18.6 |
| Перепончатокрылые (Hymenoptera) | 2 | 4.7 |
| имаго | 2 | 4.7 |
| Ichneumonidae (имаго) | 1 | 2.3 |
| Перепончатокрылые, ближе не определенные (имаго) | 1 | 2.3 |
| Скорпионницы (Mecoptera)
(личинки) |
2 | 4.7 |
| Клопы (Hemiptera) | 4 | 9.3 |
| Чешуекрылые (Lepidoptera) | 13 | 30.2 |
| имаго | 1 | 2.3 |
| личинки | 12 | 27.9 |
| Ногохвостки (Collembola) | 1 | 2.3 |
| Насекомые, ближе не определенные | 7 | 16.3 |
| имаго | 1 | 2.3 |
| личинки | 3 | 7.0 |
| куколки | 3 | 7.0 |
| Паукообразные (Arachnida) | 21 | 48.8 |
| Пауки (Araneus) | 7 | 16.3 |
| Клещи (Acarus) | 7 | 16.3 |
| Сенокосцы (Phalangium) | 13 | 30.2 |
| Многоножки (Myriapoda)
Chilopoda |
4 | 9.3 |
| Моллюски (Mollusca) | 2 | 4.7 |
| Малощетинковые черви (Lumbricidae) | 27 | 62.8 |
| Растительный материал | 3 | 7.0 |
| Вегетативные органы | 1 | 2.3 |
| Семена | 1 | 2.3 |
| Число желудков с кормом | 43 | 100 |
| Число пустых желудков | 1 | 2.3 |
Размножение и экологическая структура популяции
Для западной (европейской) части ареала этого вида утвердилось мнение, что начало воспроизводства у него сдвинуто здесь на более поздний срок (примерно на месяц) по сравнению с другими, симпатрическими видами бурозубок, например S. аraneus. По наблюдению же сибирских исследователей (Ревин, 1968; Юдин, 1971, Докучаев, 1990), в восточных частях ареала продолжительность размножения и его интенсивность у равнозубой бурозубки в общем сходны с таковыми других представителей рода Sorex; рождение молодых и выход их из гнезд приурочены к тем же срокам, но несколько отодвинуты к осени. Точными сведениями о сроках начала размножения равнозубой бурозубки в нашем регионе мы не располагаем, но, судя по тому, что все три случая поимки здесь беременных самок (16.08.1969, 7.07.1978 и 23.07.1989) пришлись на вторую половину лета, кормящей – на начало августа (6.08.1990), а первых прибылых – на конец июня (25.06.1999 и 27.06.2002), вступление местной популяции в воспроизводство следует относить к началу мая, разгар его – к концу июля, а завершение – на сентябрь.
В Северо-Восточном Забайкалье первый этап размножения, по-видимому, проходит на 10–15 дней раньше: беременные самки начинали попадаться в III декаде апреля, а самостоятельный молодняк – в I–II декадах мая (Лямкин и др., 1985). В Северном Приохотье, по данным Н. Е. Докучаева (1990), спаривание у самок равнозубой бурозубки происходит в основном в третьей декаде мая, о чем свидетельствуют даты отлова первых сеголеток. Молодые зверьки равнозубой бурозубки в уловах за разные годы отмечались там в следующие сроки: 1981 г. – 2.07.1982 г. – 9.07.1982 г. –15.07.1984 г. – 10.07.1985 г. – 4.07.1986 г. – 12.07.1987 г. – 29.06.1988.
Принимая время между спариваниями и выхода из гнезд молодняка, равным 42 суткам (Skaren, 1979), получим сроки спаривания, указанные выше. Самое раннее спаривание приходилось на 18 мая.
Второй раз самки спариваются сразу же после рождения первого выводка. Так, 27.06.1961 были добыты две кормящие самки, у которых в матках просматривались эмбрионы следующей беременности. По времени второе спаривание приходилось на конец июня – начало июля. Со второй декады июля уже отмечались самки, кормящие второй помет. Сроки третьей беременности у самок равнозубой бурозубки сильно размыты, и самки, беременные третьим выводком, ловились с последних чисел июля. В основном же третья беременность проходила в августе или в сентябре. Так, по свидетельстиву Н. Е. Докучаева (1990), 27 августа 1983 г. была добыта кормящая самка, у которой можно было различить послеплодные пятна трех беременностей.
В сентябре размножение заканчивалось, но у части самок оно затягивалось до октября. К примеру, 21.09.1979 г. была отловлена недавно родившая самка с хорошо развитыми млечными железами, а 16.09.1981 – самка на поздних стадиях беременности. Эти самки смогли закончить кормление своих детенышей в лучшем случае в октябре. Таким образом, общая продолжительность периода размножения у равнозубых бурозубок Северного Приохотья составляет 4.5 месяца.
У равнозубых бурозубок по сравнению с другими видами рода Sorex сроки окончания размножения смещены на более позднее время, на что впервые обратил внимание Б. С. Юдин (1971). Позднее пролонгированный характер воспроизводства этой бурозубки был подтвержден в двух удаленных друг от друга регионах – нами в Карелии (Ивантер, 1975) и Лямкиным с соавторами (1985) в Северо-Восточном Забайкалье. С этим выводом согласуются и данные по Южной Якутии (Ревин, 1989), средней тайге бассейна р. Вычегды (Куприянова, 1978), а также Западному Саяну (Юдин и др., 1979). Большинство других исследователей приводят сроки размножения равнозубой бурозубки, близкие к указанным выше. Например, в Амурской области (Бромлей и др., 1984) период размножения длится с конца мая по сентябрь включительно, т. е. в те же сроки, которые указаны выше. Остальные 4 взрослые самки были пойманы уже после окончания размножения – 8 и 27 сентября, 9 и 29 октября.
Матка взрослых самок S. isodon крупная (суммарная длина рогов 14.4–34.0, в среднем 26.1 мм, тела 7.3–11.4, в среднем 8.5 мм), с относительно удлиненными рогами. Индекс, т. е. отношение длины тела матки к суммарной длине рогов, равен 32.4 %. У молодых самок этот индекс такой же – 33 %, средняя длина тела матки составляет у них 4.3 (3.5–6.5) мм, рогов 12.7 (9.3–15.5) мм.
Размеры и гистологическое строение семенников зимовавших самцов (добытых 27.05, 18.06 и 9.07.1967 и 19.08.1971) свидетельствуют об активном сперматогенезе (рис. 2). Длина семенников варьирует от 7.2 до 8.1 мм, равняясь в среднем 7.8 мм, длина семенных пузырьков 8–10.5 (в среднем 8.8) мм, вес двух семенников 190–322 (в среднем 238) мг. У молодых неполовозрелых самцов (n = 21) длина семенников 1–2 (в среднем 1.3) мм, вес 1–3 (в среднем 2.1) мг. Гистологическая картина гонад этих самцов свидетельствует о половом покое.
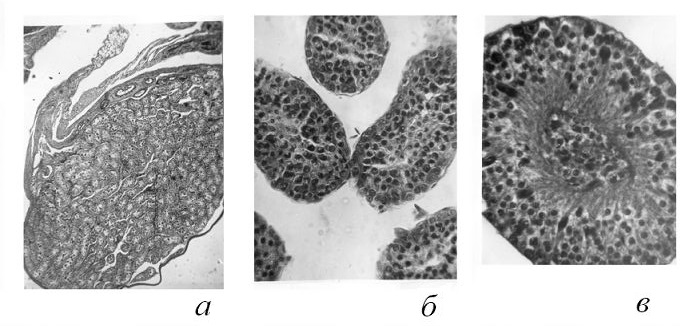
Рис. 2. Гистологическая структура семенников равнозубой бурозубки: а – в состоянии покоя, б – пассивного, в – активного сперматогенеза
Fig. 2. Histologic structure of the testicles of an equal-toothed shrew. a – at rest, б – in passive state, в – in active state
В Карелии молодые особи ловились начиная с 9 июля (1959 г.). Судя по этой дате, рождение их приурочено к середине июня. Таким образом, размножение у равнозубой бурозубки в условиях северо-западной периферии ареала не только заканчивается, но и начинается позднее, чем у других видов Soricidae. При этом в размножении участвуют преимущественно взрослые, зимовавшие самки, и общая доля созревающих в год рождения прибылых не превышает 6–8 %.
Размер выводка у равнозубой бурозубки Карелии, по данным обследования беременных зверьков, находился в пределах от 2 до 11 эмбрионов на самку (в среднем 7.7 + 0.3). В Сибири же, согласно данным Н. Е. Докучаева (1990), плодовитость у самок этого вида снижалась от июня к сентябрю и от первой беременности к третьей (табл. 3).
Средняя плодовитость у равнозубой бурозубки на всем протяжении области ее распространения оказалась довольно стабильной (Докучаев, 1990). Так, на Камчатке и о-ве Парамушир на самку в среднем приходилось 7.4 эмбриона, в Приморскорм крае – 7.2, в окрестностях оз. Байкал – 7.1, на Алтае и Саянах – 7.6, в Финляндии – 7.1. Лишь в Республике Коми этот показатель был ниже – 6.1. Меньше он был и там, где материалы были получены во второй половине периода размножения, когда плодовитость у самок сильно снижена, например в Амурской области (5.0), Барабе (6.7) и т. д.
Среди изученных сибирских популяций равнозубой бурозубки участие прибылых самок в размножении в год рождения – явление довольно обычное (Докучаев, 1990). Правда, это характерно далеко не для всех представительниц этой возрастной группы, обычно в размножении принимают участие от 4–6 до 15–20 %. Лишь по данным Лямкина с соавторами (1985), в размножении в Муйской котловине (район оз. Байкал) участие молодых самок в размножении в популяции равнозубой бурозубки достигало в отдельные годы 42.4 %.
Tаблица 3. Изменения плодовитости зимовавших самок равнозубой бурозубки на реке Челомджа по месяцам и беременностям (1981–1987 гг.) (по: Докучаев, 1990)
Table 3. Changes in fertility of females of the equal-toothed shrews wintering on the Chelomdzha River on monthes and pregnancies (1981–1987) (on: Dokuchaev, 1990)
| Месяц, беременность | Общее кол-во самок | Количество самок (экз.) с данным размером выводка | Среднее число эмбрионов на самку (M ± m) | Коэффициент вариации, Cv | ||||||||||||||
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |||||||||
| Июнь | 7 | — | — | — | По месяцам | 6 | 1 | — | 9.1 ± 0.14 | 4.2 | ||||||||
| Июль | 10 | — | — | — | — | — | — | 5 | 2 | 2 | 1 | 8.9 ± 0.35 | 12.4 | |||||
| Август | 14 | 1 | 1 | — | 1 | 3 | 1 | 4 | 3 | — | — | 6.7 ± 0.59 | 32.8 | |||||
| Сентябрь | 4 | — | 1 | — | — | 2 | 1 | — | — | — | — | 5.5 ± 0.87 | 31.5 | |||||
| По беременностям | ||||||||||||||||||
| Первая | 5 | — | — | — | — | — | — | — | 4 | 1 | — | 9.2 ± 0.20 | 4.9 | |||||
| Вторая | 11 | — | — | — | — | — | — | 4 | 4 | 2 | 1 | 9.0 ± 0.30 | 11.1 | |||||
| Третья | 19 | 1 | 2 | — | 1 | 5 | 2 | 5 | 3 | — | — | 6.5 ± 0.48 | 32.2 | |||||
| Всего | 35 | 1 | 2 | — | 1 | 5 | 2 | 9 | 11 | 3 | 1 | 7.7 ± 0.35 | 26.9 | |||||
В Северном Приохотье Н. Е. Докучаев (1990) зафиксировал эмбриональную смертность у 35.3 % беременных самок равнозубой бурозубки. Получается, что в результате аутолиза популяция этого вида не досчиталась 7.1 % приплода. Гибель же на стадиях яйцеклеток постигает в среднем одну самку из четырех (при отходе яйцеклеток, составившем 2.8 %). В то же время из пяти случаев резорбции эмбрионов лишь один пришелся на вторую беременнсть. При этом гибель зародышей в основном отмечалась в правом роге матки. Из 10 резорбирующихся эмбрионов лишь 2 были из левого рога.
Возрастная структура популяции этой землеройки отличается относительно большим удельным весом взрослых животных, составляющих в наших сборах 22 %. Это говорит о низкой интенсивности размножения вида. В июле половой состав молодых зверьков характеризуется значительным преобладанием самцов; среди добытых в Карелии 32 сеголеток их оказалось 26 (81.2 %). У взрослых особей соотношение полов близко к 1:1.
Линька
У этого вида весенняя линька проходит, по всей видимости, в те же сроки, что и у обыкновенной бурозубки (Ивантер и др., 1985). У взрослого самца, добытого 27.05.1967, шкурка имела на 25 % площади темную мездру и рост нового волоса происходил на задней части спины. В последующие месяцы у зимовавших особей этого вида отмечается старческая линька. Ее следы в виде темных пятен на мездре, занимающих от 2 до 50 % площади шкурки и всегда расположенных на спинной стороне, обнаружены у всех добытых в июле – октябре взрослых зверьков.
Молодые линяют довольно поздно, начиная в конце сентября – октябре. Из 11 исследованных за этот период сеголеток только три (добытые 20.09.1987, 1.10.1968 и 20.10.1969) не имели следов линьки, остальные же интенсивно линяли (темные пятна на мездре занимали от 10 до 100 % площади шкурки), а с последней декады октября все отлавливавшиеся зверьки этого вида имели уже зимний мех. Правда, в литературе имеются указания и на более ранние сроки осенней линьки у сеголеток равнозубой бурозубки: 20 августа – 3 октября (Тавровский и др., 1971).
Топография же весенней линьки у этого вида несколько отличается от других. У самок она начинается на передней части тела, когда пигментацией охвачена голова, кожа вокруг передних ног и нижняя часть тела от горла до живота. Вокруг боковых желез также наблюдается рост нового волоса. У самцов линька тоже начинается на голове, затем переходит на область боковых желез, а иногда и на передние лапы. С 20-х чисел июня все зверьки были уже с коротким летним мехом. Относительно небольшой материал (9 шкурок) не позволяет более детально проследить порядок смены меха у самцов. Однако судя по наличию промежуточного весеннего меха, можно, наверное, говорить, что и у этого вида имеется две весенних линьки.
Материала по линьке самок у нас еще меньше. Три самки, отловленные 4.04 и 11.04.2001 г. и 20.04.1959, имели небольшие пятна пигмента – в основном на голове, шее и середине спины, а у самок, отловленных в мае, весенний мех наблюдался на голове и спине. Смена меха у них шла по характерному волновому типу. В летний период у перезимовавших зверьков наблюдалась старческая линька, причем у самок имелся пигмент вокруг сосков, где на месте вытертых старых волос формировались новые.
В общем смена летнего волоса на зимний идет у равнозубой бурозубки по той же схеме, что и у средней. Это, возможно, объясняется их близким филогенетическим родством (Козловский, Орлов, 1971). Линька, по Н. Е. Докучаеву (1990), начинается на спинной стороне тела, затем распространяется на голову и бока. На нижней стороне тела пигмент начинает появляться в районе грудного отдела, а затем на животе. В последнюю очередь линяют участки у передних ног и в паху, там же дольше всего сохраняется пигмент в коже. Каких-либо существенных отличий в ходе линьки у равнозубой бурозубки с Северного Приохотья (Докучаев, 1990) и Финляндии (Skaren, 1979) не обнаружено, но что касается сроков, то в бассейне реки Челомджа (Северо-Восточная Азия) зверьки начинали линять примерно на две недели раньше. Так, особи с признаками линьки регистрировались здесь с 26 августа, а полная смена летнего меха на зимний отмечена 20 сентября. В Финляндии эти сроки приходятся соответственно на 7 сентября и на 4 октября (Skaren, 1979).
По сравнению со средней бурозубкой у рассматриваемого вида осенняя линька начинается в более поздние сроки. Так, если у первого вида на Челомдже линяющие зверьки отмечались с 31 июля, то у равнозубой бурозубки – лишь с 26 августа (табл. 4), в то время как у средней бурозубки в этом месяце, как мы помним, всего 24.4 % линяющих зверьков.
В процессе осенней линьки прибылые особи равнозубой бурозубки получают зимний мех, длина которого в 1.6 раза больше летнего (табл. 5). Что же касается летнего меха у перезимовавших зверьков, то тут обнаруживается та же закономерность, что и у средних бурозубок. У этих двух видов перезимовавшие самки в летний период характеризуются более длинным мехом, чем самцы этой же возрастной группы.
Таблица 4. Интенсивность осенней линьки у сеголеток равнозубой бурозубки бассейна р. Челомджа в разные годы (самцы и самки вместе) (по: Докучаев, 1990)
Table 4. Intensity of autumn shedding of this-year broods of equal-toothed shrews in the Chelomdzha river basin in different years (all males and females together) (on: Dockuchaev, 1990)
| Год | Август | Сентябрь | ||||
| n | с линькой | n | с линькой | |||
| экз. | % | экз. | % | |||
| 1979 | Нет данных | 15 | 11 | 73.3 | ||
| 1981 | 41 | 8 | 19.5 | 20 | 13 | 65.0 |
| 1982 | 7 | 3 | 42.9 | Нет данных | ||
| 1983 | 8 | — | — | 6 | 1 | 16.7 |
| 1985 | 33 | 1 | 3.0 | 4 | 3 | 76.0 |
| 1986 | 6 | — | — | 7 | 4 | 67.1 |
| 1987 | 11 | — | — | 3 | 1 | 33.3 |
| Всего | 114 | 12 | 10.5 | 55 | 33 | 60.0 |
Н. Е. Докучаев (1990) сообщает всего о двух добытых им экземплярах равнозубой бурозубки с признаками весенней линьки. У самца, пойманного 31.03.1986, проходила первая весенняя линька, пигмент простирался по всей нижней стороне тела и захватывал бока и голову. На спине же, где еще оставался зимний мех, мездра была чистая. На боковых железах также не было пигмента, но здесь смена волос уже прошла. Самец же, отловленный 14.06.1987, находился в стадии второй весенней линьки. Пигмент у него покрывал всю поверхность кожи со стороны мездры, но на брюшной стороне тела он только начал закладываться, и эта часть шкурки была окрашена менее интенсивно. По сравнению со средней бурозубкой это были довольно поздние сроки второй весенней линьки.
Таблица 5. Сезонная изменчивость высоты меха (мм) у различных половозрелых групп равнозубой бурозубки челомджинской популяции (по: Докучаев, 1990)
Table 5. Seasonal variability of the fur height (mm) in different mature groups of the Chelomdzha popoulation of equal-toothed shrews (on: Dockuchaev, 1990)
| Возраст, пол | Зимний мех | Весенний мех | Летний мех | |||
| M ± m | M ± m | M ± m | ||||
| Зимовавшие | ||||||
| самцы | 5 | 8.1 ± 0.20 | 3 | 6.4 ± 0.12 | 19 | 4.0 ± 0.06 |
| самки | 2 | 7.8 ± 0.10 | — | — | 18 | 4.5 ± 0.05 |
| сеглетки (самцы и самки вместе) | 3 | 7.8 ± 0.17 | — | — | 70 | 4.9 ± 0.03 |
Заключение
Как показали исследования, выявленные выше экологические особенности, свойственные равнозубой бурозубке в специфических условиях северо-западной периферии ареала, отличаются значительной лабильностью и гармоничной согласованностью как с плотностью и состоянием популяций, так и с внешними факторами среды. Разнообразие экологических приспособлений, их трансформация и смена во времени и пространстве, адаптивный динамизм и ориентированность на конкретную экологическую ситуацию, складывающуюся из взаимодействия экзогенных и эндогенных факторов, обуславливают относительную стабильность и оптимальное состояние популяций рассматриваемого вида в экстремальных для него приграничных условиях. Вместо узких специальных приспособлений индивидуального ранга в периферических его населениях успешно реализуются широкие структурно-популяционные адаптивные комплексы, отличающиеся динамичностью и высокой скоростью компенсаторной перестройки. Определенную роль в жизни и особенно воспроизводстве периферических популяций вида играют специфические регуляторные механизмы направленного действия, осуществляющие поддержание численности на уровне усовершенствованной групповой организации. Однако в северных границ ареала действие этих механизмов более ограничено и поддержание динамичного баланса популяции с ресурсами ее местообитания в основном достигается при значительном участии внешних факторов.
Библиография
Бобрецов А. В., Нейфельдт Н. Д., Сокольский Л. М. и др. Млекопитающие Печоро-Илычского заповедника . Сыктывкар: Коми книжн. изд-во, 2004. 464 с.
Большаков В. Н., Васильев А. Г., Шарова Л. П. Фауна и популяционная экология землероек Урала (Mammalia. Soricidae) . Екатеринбург, 1996. 268 c.
Бромлей Г. Ф., Костенко В. Я., Николаев И. П. и др. Млекопитающие Зейского заповедника . Владивосток: ДВНЦ АН СССР, 1984. 139 с.
Вольперт Я. Л. Размножение бурозубок (род Sorex) Северо-Восточной Якутии // Охотничьи и промысловые ресурсы Сибири. Новосибирск: Наука, 1986. С. 209–218.
Вольперт Я. Л., Аверенский А. И. Характеристика питания землероек-бурозубок рода Sorex в Западном Предверхоянье // Териологические исследования в Якутии. Якутск, 1983. С. 57–73.
Докучаев Н. Е. Структура и продуктивность сообществ землероек-бурозубок (Insectivora, Soricidae) Чукотки // Зоологический журнал. 1994. Т. 73 (9). С. 114–123.
Докучаев Н. Е. Экология бурозубок Северо-Восточной Азии . М.: Наука, 1990. 160 с.
Докучаев Н. Е. Питание землероек-бурозубок (Soricidae) и оценка их роли в горно-таежных экосистемах Северо-Восточной Сибири // Экология млекопитающих Северо-Восточной Сибири. М.: Наука, 1981. С. 3–22.
Долгов А. А. Бурозубки Старого Света . М.: Изд-во МГУ, 1985. 219 с.
Долгов В. А., Чабовский В. И., Шилова С. А., Эфрон К. М. Некоторые вопросы экологии бурозубок и их значение в очагах клещевого энцефалита // Бюллетень Московского общества испытателей природы. Отд. биол. 1968. Т. 73. Вып. 6. С. 17–28.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР . Л.: Наука, 1975. 246 с.
Ивантер Э. В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала . М.: Товарищество научных изданий КМК, 2018. 770 с.
Ивантер Э. В., Ивантер Т. В. К изучению равнозубой бурозубки (Sorex isodon Turov, 1924) // Фауна и экология наземных позвоночных. Петрозаводск, 1988. С. 88–112.
Ивантер Э. В., Ивантер Т. В., Лобкова М. П. О питании землероек-бурозубок (Sorex L.) Карелии // Труды государственного заповедника «Кивач». Петрозаводск, 1973. Вып. 2. С. 154–169.
Ивантер Э. В., Ивантер Т. В., Туманов И. Л. Адаптивные особенности мелких млекопитающих . Л.: Наука, 1985. 318 с.
Ивантер Э. В., Макаров А. М. Территориальная экология землероек-бурозубок (Insectivora, Sorex) . Петрозаводск: Изд-во Петрозавод. гос. ун-та, 2001. 272 с.
Карасева Е. В., Телицына А. Ю., Жигальский О. А. Методы изучения грызунов в полевых условиях . М.: Из-во ЛКИ, 2008. 416 с.
Козловский А. И., Орлов В. Н. Кариологическое подтверждение видовой самостоятельности равнозубой бурозубки Sorex isodon Turov (Insectivora, Soricidae) // Зоологический журнал. 1971. Т. 50. Вып. 7. С. 1056–1061.
Куприянова И. Ф. Биотопические группировки мелких млекопитающих и динамика их численных взаимоотношений на юге Архангельской области // Фауна и экология позвоночных животных. М.: Наука, 1978. С. 114–130.
Куприянова И. Ф. Сем. Soricidae – землеройковые // Фауна европейского Северо-Востока России. Млекопитающие. СПб.: Наука, 1994. Т. 2. Ч. 1. С. 11–67.
Куприянова И. Ф., Ануфриев В. М. Размещение бурозубок в естественных и антропогенных местообитаниях средней тайги Коми АССР // I Всесоюзное совещание по биологии насекомоядных млекопитающих. М., 1992. С. 89–91.
Лукьянова И. В. Растительные корма в пищевом рационе землероек (Soricidae) // Териология. Т. 2. Новосибирск: Наука, 1974. С. 281–283.
Лукьянова И. В. Распределение землероек (Soricidae) в южной тайге и подтаежных лесах Приобья // Систематика, фауна, зоогеография млекопитающих и их паразитов. Новосибирск, 1975. С. 70–76.
Лукьянова И. В. Особенности распространения равнозубой бурозубки (Sorex isodon Turov) в Западной и Средней Сибири // I Всесоюзное совещание по биологии насекомоядных млекопитающих. М., 1992. С. 103–105.
Лямкин В. Ф., Пузанов В. М., Малышев Ю. С. Некоторые особенности размножения бурозубок (род Sorex) Северного Забайкалья // Фауна и экология млекопитающих Якутии. Якутск: Изд. ЯФ СО АН СССР, 1985. С. 73–84.
Макаров А. М. К территориальному поведению равнозубой бурозубки (Sorex isodon Turov) // Животный мир Енисейской тайги и лесотундры и природная зональность. М., 1983. С. 215–230.
Моралева Н. В. К проблеме межвидовых отношений близких видов землероек-бурозубок (Insectivora. Sorex) // Фауна и экология птиц и млекопитающих Средней Сибири. М.: Наука, 1987. С. 213–228.
Нестеренко В. А. Насекомоядные юга Дальнего Востока и их сообщества . Владивосток: Дальнаука, 1999. 173 с.
Новиков Г. А. Полевые исследования экологии наземных позвоночных . Л.: Сов. наука, 1953. 602 с.
Охотина М. В. Морфоэкологические особенности различных видов бурозубок (Sorex, Insectivora), обуславливающие возможность их совместного существования // Фауна и экология наземных позоночных юга Дальнего Востока СССР: Тр. биолого-почвенного ин-та. Владивосток, 1974. Т. 17. С. 42–57.
Попов М. В. Средняя, малая, равнозубая бурозубки // Млекопитающие Якутии. М.: Наука, 1971. С. 57–71.
Пучковский С. В. Опыт сравнительного изучения представителей двух стратегий выживания : Автореф. дис. ... д-ра биол. наук. Петрозаводск, 2000. 40 с.
Пучковский С. В. К вопросу о закономерностях биотопического распределения бурозубок (Sorex) // Научные труды Свердловского государственного педагогического института. 1973. Сб. 221. С. 109–125.
Пучковский С. В. Биотопические особенности веса тела и некоторых внутренних органов бурозубок (Sorex, Soricidae, Insectivora) // Научные доклады высшей школы. Сер. биол. науки. 1969. № 9. С. 43–48.
Равкин Ю. С., Лукьянова И. В. География позвоночных южной тайги Западной Сибири . Новосибирск, 1976. 338 с.
Ревин Ю. В. Эколого-фаунистический очерк насекомоядных и мелких грызунов Олекмо-Чарского нагорья // Материалы по биологии и динамике численности мелких млекопитающих Якутии. Якутск, 1968. С. 5–86.
Тавровский В. А., Егоров А. В., Кривошеев В. Г. и др. Млекопитающие Якутии . М.: Наука, 1971. 670 с.
Шварц Е. А., Демин Д. В. О факторах сосуществования родственных видов в местах симпатрии ареалов (на примере Soricidae) // Доклады АН СССР. 1986. Т. 289. № 1. С. 255–256.
Юдин Б. С. Фауна землероек (Mammalia. Soricidae) севера Западной Сибири // Биологические проблемы Севера. Магадан, 1971. Вып. 48. С. 48–53.
Юдин Б. С., Барсова Л. И. Землеройки кедровых лесов Прителецкого очага клещевого энцефалита // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука, 1967. С. 51–59.
Юдин Б. С., Галкина Л. И., Потапкина А. Ф. Млекопитающие Алтае-Саянской страны . Новосибирск: Наука. Сиб. отд-ние, 1979. 293 с.
Hanski I. Food consumption, assimilation and metabolic rate in six species of shrews (Sorex and Neomys) // Ann. Zool. Fennici. 1984. Vol. 21. P. 157–165.
Hanski I., Kaikusalo A. Distribution and habitat selection of shrews in Finlkand // Ann. Zool. Fennici. 1989. Vol. 26. No 4. P. 339–348.
Siivonen L. Sorex isodon Nurov (1924) and S. ungulates Dobson (1890) as independent shrew species // Aquilo. Ser. Zool. 1965. Vol. 4. P. 3–49.
Skaren U. Variation, breeding and moulting in Sorex isodon Turov in Finland // Acta zool. fenn. 1979. Vol. 159. P. 1–30.
Skaren U. Variation in two shrews, Sorex unguiculatus Dobson and S. a. araneus L. // Ann. Zool. Fenn. 1964. Vol. 1. P. 2.

 © 2011 - 2026
© 2011 - 2026