



Введение
Внутривидовая и межвидовая конкуренция, ее роль в регуляции численности и эволюции разных видов принадлежит к числу тем, обсуждаемых со времен Чарльза Дарвина. В ХХ столетии в научный оборот вошло понятие «экологическая ниша», которое легло в основу концепции, связавшей представления о конкурентных отношениях и распространении видов в природе, процессах естественного отбора и видообразования, в разработку которой внесли вклад многие исследователи. К настоящему времени она вошла в круг теорий, центральных в изучении экосистем и эволюции видов, и часто рассматривается как фундаментальная для экологии в целом (Пианка, 1981; Хлебосолов, 1999; 2002; Розенберг, Рянский, 2005 и др.).
Базовым постулатом теории экологической ниши является принцип конкурентного исключения Гаузе (Gause, 1934, цит. по: Одум, 1986), согласно которому ниши совместно обитающих видов могут перекрываться лишь частично. Отношения между такими видами находятся в динамическом равновесии, которое при стабильных условиях относительно устойчиво, но при изменении ситуации нарушается и может сдвигаться в пользу одного из них. В природе одновременно действует большое количество факторов, что затрудняет оценку влияния каждого из них на популяционные процессы и динамику численности отдельных видов. Поэтому, несмотря на достаточно глубокую теоретическую проработку проблемы, пока «не удается решить вопрос о роли конкуренции в расхождении организмов по экологическим нишам и формировании сообществ» (Хлебосолов, 2002, стр. 1019).
К основным объектам конкуренции у птиц, наряду с кормовыми ресурсами, относятся места для размещения гнезд (Newton, 1998). В небольшую группу видов, позволяющих напрямую наблюдать конкурентную борьбу за места для гнездования, входят виды, использующие готовые дупла, – вторичные дуплогнездники, репродукцию которых можно контролировать, привлекая их в искусственные гнездовья. Увеличение их численности при развеске дуплянок рассматривают как прямое доказательство того, что ее ограничителем является нехватка естественных дупел (Haartman, 1971; Newton, 1994).
Борьба за места для размещения гнезд между вторичными дуплогнездниками происходит в разных формах, вплоть до убийства конкурентов, что отражается на успешности размножения, гнездовой плотности населения (Haartman, 1971) и даже возможностях гибридизации конкурирующих видов (Vallin et al., 2012). При этом нередко птицы, демонстрирующие крайне конфликтное поведение в одних случаях, не обнаруживают явных признаков агрессивности в других ситуациях и регионах (Creutz 1955. цит. по Slagsvold, 1975; Haartman, 1957, 1971; Tompa, 1967; Slagsvold, 1975, 1978; Minot, Perrins, 1986; Merilä, Wiggins, 1995; Krist, 2004; Смирнов, 2012 и др.). Степень напряженности межвидовых отношений и ее влияния на популяционные характеристики связывают с особенностями местообитаний, различиями в обилии дупел, численности птиц, времени откладки яиц (Slagsvold, 1975; Ahola et al., 2007 и др.). При изучении этих зависимостей, как правило, используют данные, полученные в разных регионах или на территориях с непохожими экологическими условиями. Однако результаты таких сравнений редко бывают однозначными из-за невозможности учесть влияние большого количества факторов, трудно или вовсе не поддающихся оценке. Еще более ограничены возможности оценки их комплексного влияния на межвидовые отношения птиц и анализа всех аспектов работы конкурентных механизмов в естественной среде. По этим причинам многие моменты, связанные с межвидовой конкуренцией у птиц, нуждаются в уточнении и проверке в разных условиях.
Для климата Карелии характерна широкая амплитуда межгодовых колебаний сезонных температур, благодаря чему фенологическая обстановка весеннего периода может существенно отличаться даже в смежные годы. Эти колебания по-разному отражаются на сроках прилета и размножения отдельных видов птиц, что приводит к ежегодным изменениям структуры их межвидовых отношений. Сравнительный анализ данных, полученных в Карелии в 1975 г. с беспрецедентно ранней весной и в 1976 г. с поздней весной, позволил отследить изменения напряженности конкурентных отношений дуплогнездников при резких межгодовых перепадах температур предбрачного периода и оценить последствия этих колебаний для воспроизводства их северных популяций.
Материалы
Проанализированы данные, собранные в Кижских шхерах Онежского озера (62° с. ш.) в 1975–1977 гг. с разным характером погодных условий предгнездового периода (табл. 1). Весна 1975 г. была необычайно ранней и устойчиво теплой: с середины марта среднедекадные температуры не опускались ниже 0° (Скороходова, 2008); снег в лесах и лед на Онежском озере сошли к середине апреля. Весна 1976 г. выдалась поздней и холодной, лето – дождливым, а осень и зима – ранними. Температуры 1977 г. были близки к средним многолетним для региона.
Таблица 1. Характеристика весенних условий 1975–1977 гг. (метеостанция на острове Маячный, Онежское озеро)
Table 1. Characteristics of spring conditions 1975–1977 (weather station on the island Mayachny, Lake Onega)
| Год | Переход min t° С через 0° |
Переход min t °С через +3° | Начало навигации | Средние min t °C в данные декады мая | Даты последнего похолодания (ниже +3°) | ||
| I | II | III | |||||
| 1975 | 20.04 | 8.05 | 18.04 | 2.7 | 4.8 | 3.3 | 26–30.05 (снегопады) |
| 1976 | 4.05 | 18.05 | 23.05 | –1.5 | 2.5 | 2.1 | 21–30.05 |
| 1977 | 30.04 | 14.05 | ?.05 | 0.1 | 3.4 | 2.4 | 21–30.05 |
В конце марта – начале апреля 1975 г. в полуоткрытых стациях, лиственных и смешанных лесах о–ва Клименецкий (147 км²) на четырех маршрутах общей протяженностью 10 км через каждые 50–70 м были развешены искусственные гнездовья – дощатые синичники с диаметром летка 38–40 мм. Такие размеры летка не позволяли проникнуть в них дятлам или мелким хищникам, поэтому основные потери из-за разорения гнезд были связаны с деятельностью птиц- конкурентов. Ежегодно от 33 до 60 % гнездовий занимали птицы четырех видов (табл. 2).
Таблица 2. Заселенность искусственных гнездовий на о-ве Клименецком в 1975–1977 гг.
Table 2. The number of bird boxes used by secondary hole-nesting birds on the island Klimenetsky in 1975–1977
| Показатели | 1975 г. | 1976 г. | 1977 г. | Всего | ||||
| N | % | n | % | N | % | n | % | |
| Всего синичников, | 196 | 100 | 188 | 100 | 167 | 100 | 551 | 100 |
| из них занято | 86 | 43.9 | 112 | 59.6 | 55 | 32.9 | 253 | 45.7 |
| Всего гнезд с кладкой*, в т. ч. | 88 | 100 | 121 | 100 | 55 | 100 | 264 | 100 |
| P. major | 32 | 36.4 | 37 | 30.6 | 8 | 14.6 | 77 | 29.2 |
| F. hypoleuca | 54 | 61.3 | 72 | 59.5 | 45 | 81.8 | 171 | 64.7 |
| Ph. phoenicurus | 2 | 2.3 | 7 | 5.8 | 1 | 1.8 | 10 | 3.8 |
| J. torquilla | – | – | 5 | 4.1 | 1 | 1.8 | 6 | 2.3 |
* – в некоторых синичниках птицы гнездились дважды за сезон.
Методы
Все искусственные гнездовья осматривали один раз в 5–7 дней с середины апреля до конца июля в 1975 г. и до середины июля в 1976 г., а также один раз в 10–14 дней в течение июня – июля 1977 г.
Птиц отлавливали и кольцевали на гнездах, в основном, перед вылетом птенцов. Вторыми кладками больших синиц, которых не удалось пометить при первом гнездовании, считали кладки, появившиеся в соответствующие сроки в ближайших дуплянках. Ошибка маловероятна из-за значительных расстояний между участками соседних пар.
Численность птиц в естественных биотопах, приведенная в видовых очерках, указана по данным учетов на тех же маршрутах в 1974 г. до развески синичников и в 1995–2013 гг. после их полного разрушения.
При обработке данных использованы обычные методы вариационной статистики (Лакин, 1990; Ивантер, Коросов, 2003).
Результаты
В шхерах искусственные гнездовья заселяли четыре вида птиц (табл. 2), принадлежащих к разным систематическим группам. Большая синица Parus major L., мухоловка-пеструшка Ficedula hypoleuca L. и садовая горихвостка Phoenicurus phoenicurus (L.) относятся к разным семействам отряда Воробьинообразных Passeriformes, вертишейка Jynx torquilla L.– мелкий представитель отряда дятлообразных Piciformes. Эти виды, занимающие каждый свою специфическую экологическую нишу и отличающиеся друг от друга по большинству параметров (численности, характеру и срокам пребывания в регионе, особенностям питания и биологии и пр.), используют для гнездования один общий ресурс – готовые дупла, которые не могут выдалбливать самостоятельно.
Большая синица Parus major L. – вид, гнездящийся и зимующий в регионе. Встречается на всех островах Кижских шхер, покрытых лесом. Минимальная величина острова, где в естественном дупле низкой березы на высоте 1 м найдено гнездо, – 30 м х 80 м. Избегает участков с преобладанием хвойных пород. В предпочитаемых стациях (приопушечная полоса лиственных лесов и куртины деревьев среди лугов) численность составляет 1–3 пары/км², на участках с развешенными синичниками – до 25 пар/км². Осуществляет в Карелии два цикла размножения.
Птицы, зимующие у жилья человека, появляются в лесах островов в конце марта и первое время кочуют со стайками пухляков. Распределение по участкам заканчивается к середине апреля. В 1975 и 1976 гг. в дуплянках гнездилось по 16 пар. В 1975 г. массовая постройка гнезд проходила в 20-х числах апреля, первые яйца отложены 30.04–4.05, выводки покинули гнезда 12–18.06. Еще до их вылета родители начинали носить мох в соседние дуплянки и за день (2 случая), в день вылета или на следующий после него приступали ко вторым кладкам. Поздней весной 1976 г. размножение началось на 2 недели позже: постройка гнезд – с 5.05, откладка яиц – с 10.05, кладки второго цикла начаты только через 5–8 дней после вылета первых выводков (табл. 3).
Несмотря на сдвиг сроков, плодовитость синиц в 1976 г. осталась на прежнем уровне. Кладки по величине не уступали прошлогодним, и все пары с успешным первым циклом размножения начали второй, в четырех случаях – обновив гнезда в тех же синичниках. Еще в двух домиках синицы построили гнезда поверх 2 яиц вертишейки и кладки мухоловки-пеструшки. Вместе с тем успешность гнездования снизилась. Из-за неблагоприятных условий увеличился частичный отход яиц (неоплодотворенные и с погибшими эмбрионами) и слабых птенцов, что достоверно (T = 2.86, p > 0.99, df = 23) сократило величину даже майских выводков (проследить до конца историю всех поздних гнезд, к сожалению, не удалось). Еще более заметный ущерб нанесли конкуренты. Вертишейки вытащили несколько недостроенных гнезд синиц и начатую кладку, отложив в дуплянки свои яйца. Возросло число полностью разоренных и возобновленных кладок. Это способствовало падению численности птиц, сократившейся в 1977 г. до 5 пар. Также уменьшилась средняя величина кладок (T = 3.48, p > 0.999, df = 27) и выводков (табл. 3).
Таблица 3. Показатели репродукции больших синиц P. major на о-ве Б. Клименецком в 1975–1977 гг.
Table 3. Indicators of reproduction of Great Tit P. major on the island B. Klimenetsky, 1975–1977
| Период | Показатели | 1975 | 1976 | 1977 | |
| Апрель –май | Дата начала кладок | M (n)
Lim |
4.05 (18)
30.04–20.05 |
14.05 (17)
10.05–27.05 |
23.05 (4)
17.05–28.05 |
| Число кладок (в т. ч. повторных) | начатых
успешных |
18 (2)
15 |
17 (1)
12 |
5
4 |
|
| Величина полной кладки | M ± m
Lim (n) |
12.56 ± 0.23
11–14 (16) |
12.71 ± 0.33
10–15 (14) |
9.40 ± 0.57
8–11 (5) |
|
| Величина выводка | M ± m
Lim (n) |
11.93 ± 0.35
9–13 (14) |
9.55 ± 0.78
5–13 (11) |
7.50 ± 0.71
7–8 (2) |
|
| Июнь –июль | Дата начала кладок | M (n)
Lim |
14.06 (13)
12.06–20.06 |
1.07 (18)
3.06–11.07 |
29.06 (3)
26.05–1.07 |
| Число кладок (в т. ч. повторных) | начатых
успешных |
14
13 |
20 (6)
? |
3 (1)
1 |
|
| Величина полной кладки | M ± m
Lim (n) |
11.07 ± 0.41
9–14 (13) |
10.00 ± 0.53
8–13 (9) |
10
10 (1) |
|
| Величина выводка | M ± m
Lim (n) |
10 ± 0.53
7–12 (9) |
? | 10
10 (1) |
|
| Всего | Величина полной кладки | M ± m | 11.90 ± 0.60 | 11.65 ± 0.40 | 9.50 ± 0.47 |
| Величина выводка | M ± m | 11.17 ± 0.57 | ? | 8.33 ± 1.65 | |
Мухоловка-пеструшка Ficedula hypoleuca L. – перелетный вид с моноцикличным размножением. Гнездится в шхерах в лесах разных типов, избегая только сомкнутых ельников. Самая массовая из птиц, заселяющих искусственные гнездовья, при развеске которых численность птиц выросла с 3–4 до 40–50 пар/км², превысив в отдельных оптимальных для вида участках лиственных лесов 10 пар/га.
В 1975 и 1976 гг. появление самцов зарегистрировано 5 мая, самок – 10 и 13 мая. Теплой весной самцы с первых же дней активно пели и осматривали дуплянки. Холодной весной первая песня отмечена лишь на пятый день после прилета, а интенсивное пение – после появления самок. Строительство гнезд в 1975 г. проходило раньше и быстрее, интервал между началом постройки и кладкой оказался короче – соответственно 6,5 и 8,8 дня (Хохлова, Головань, 1981). Откладка яиц началась на неделю раньше, кладки и выводки были достоверно крупнее, а результативность размножения – выше (табл. 4, 5). Снижение успешности гнездования обеспечил, в основном, высокий частичный отход во время похолодания со снегопадами в конце мая. В кладках, начатых в этот период (n = 22), число неоплодотворенных яиц доходило до 5, а одна (7 яиц) состояла из них полностью. В целом они составили 11.75 ± 0.79 % от общего числа отложенных яиц, тогда как в кладках, начатых в другое время, – 6.14 ± 0.65 %.
Таблица 4. Величина кладки мухоловки-пеструшки F. hypoleuca на о-ве Клименецком в 1975–1977 гг.
Table 4. Clutch size of Pied Flycatcher F. hypoleuca on the island Klimenetsky, 1975–1977
| Год | Число яиц в кладках, начатых в данные сроки, M ± m (число кладок, n) | ||||
| май | июнь | Всего | |||
| II | III | I | II + III + июль | ||
| 1975 | 7.5 (2) | 7.42 ± 0.12 (39) | 7.18 ± 0.24 (11) | 5.50 (2) | 7.32 ± 0.10 (54) |
| 1976 | – | 6.69 ± 0.20 (32) | 6.10 ± 0.21 (29) | 5.50 ± 0.47 (6) | 6.33 ± 0.14 (67) |
| 1977 | 6.0 (1) | 6.77 ± 0.20 (26) | 6.44 ± 0.17 (18) | – | 6.49 ± 0.15 (45) |
| всего | 7.00 + 1.07(3) | 7.00 ± 0.10 (97) | 6.41 ± 0.13 (58) | 5.50 ± 0.20 (8) | 6.73 ± 0.08 (166) |
Таблица 5. Успешность гнездования мухоловки-пеструшки F. hypoleuca в 1975–1977 гг.
Table 5. The nesting success of Pied flycatcher F. hypoleuca, 1975–1977
| Показатели | 1975 | 1976 | 1977 | Всего |
| Даты начала кладок, M (lim) | 15.05–30.06 | 23.05–1.07 | 20.05–9.06 | 15.05–1.07 |
| Прослежено яиц, из них погибли: |
390 | 420 | 244 | 1054 |
| Не оплодотворены | 33 | 36 | 13 | 82 |
| С погибшими эмбрионами | 20 | 14 | 2 | 36 |
| Разорение всей кладки | – | 34 | 13 | 47 |
| Разорение частичное (1–2 яйца) | 1 | 2 | 1 | 4 |
| Гибель самки | – | – | 7 | 7 |
| Вылупилось птенцов | 336 | 334 | 208 | 878 |
| Успешность насиживания, % | 86.2 | 79.5 | 85.3 | 83.3 |
| Прослежено птенцов, из из них погибли: | 336 | 251 | 186 | 773 |
| Слабые, в дни вылупления | 7 | 7 | 8 | 22 |
| Из-за недокорма | 2 | 8 | 3 | 13 |
| Из-за гибели самки | – | – | 7 | 7 |
| Разорено гнездо | 1 | 6 | – | 7 |
| Вылетело птенцов | 326 | 230 | 168 | 724 |
| Успешность выкармливания, % | 97.0 | 91.6 | 90.3 | 93.7 |
| Общая успешность размножения, % | 83.6 | 72.9 | 77.0 | 78.0 |
| Число слетков на самку | 6.0 | 4.4 | 5.1 | 5.1 |
Благополучное размножение в 1975 г. способствовало подъему численности птиц на следующий год (табл. 2). Но репродуктивные показатели в неблагоприятных условиях 1976 г. снизились, а успешность гнездования упала из-за участившихся случаев разорения (табл. 5). Только вертишейки уничтожили 8 полных кладок мухоловок-пеструшек, поверх еще одной построили гнездо большие синицы, два гнезда были брошены птицами из-за беспокойства при откладке яиц, и 10 – до ее начала. Не слишком успешное размножение внесло свой вклад в снижение численности птиц в 1,5 раза в следующем, 1977 г. Остальные показатели при этом улучшились, хотя и не достигли значений 1975 г.
Аналогичные межгодовые изменения популяционных параметров обнаружены в эти годы и на орнитологической станции «Гумбарицы» в восточном Приладожье, где искусственные гнездовья использовались с 1970 г. (Хохлова, Головань, 1981).
Садовая горихвостка Phoenicurus phoenicurus (L.) – перелетный вид, гнездящийся на островах, покрытых лесами разных типов. В Карелии обычно ограничивается одним репродуктивным циклом, но известны случаи «неежегодной бициклии», когда пара успевала выкормить за сезон два выводка (Зимин, Ивантер, 1969). В 1970-е годы началась депрессия численности вида, затронувшая популяции на Северо-Западе России (Зимин и др., 1993). Ее дальнейшее углубление послужило основанием для включения горихвостки в качестве уязвимого вида в Красную книгу Карелии (Хохлова, 1995). В период наблюдений гнездовая плотность птиц в Кижских шхерах характеризовалась очень низким уровнем и была подвержена большим межгодовым колебаниями. В 1975 г. горихвостки заняли только 2 из 100 синичников, развешенных на о-ве Б. Клименецком в лесных угодьях с сомкнутым древостоем, и не заинтересовалась гнездовьями в полуоткрытых стациях. Постройка гнезд была начата 13.05, откладка яиц – 18 и 23.05. Полные кладки содержали по 7 яиц, выводки – 7 и 3 слетка, поскольку в кладке, которая шла во время похолодания, 4 яйца не были оплодотворены. В 1976 г. численность выросла, и птицы заселили уже 6 искусственных гнездовий также только в лесных местообитаниях. Первая песня зарегистрирована 7.05, постройка гнезд, как и в прошлом году, началась 13.05, кладки по 5, 6, 7, 7 и 8 яиц – между 20 и 26 мая, еще одна из 4 яиц – 23.06. В последнем случае пара заняла синичник, из которого только 20.06 было вынуто гнездо мухоловки, брошенное с неполной кладкой. Из всех гнезд вылетели птенцы, но в кладке из 7 яиц одно не содержало зародыша, и в позднем гнезде два эмбриона погибли за 2–3 дня до вылупления. В 1977 г. численность птиц вновь упала, а единственное гнездо, найденное в искусственных гнездовьях, было разорено, по-видимому, вертишейкой, поселившейся на этом же участке.
Вертишейка Jynx torquilla L. – перелетный вид, обитающий в основном в лиственных лесах островов. Предпочитает разреженные фаутные древостои и куртины древесно-кустарниковой растительности среди сенокосных лугов. В 1975 г. на 1 км² таких угодий можно было встретить 2–4 пары птиц, в 1976 г. – 5–6 пар.
В 1975 г. первые птицы зарегистрированы 8.05, однако в синичниках они не поселялись и их не разоряли, хотя рядом найдены 2 гнезда, устроенных в старых дуплах больших пестрых дятлов в осине (на высоте 5 м) и трухлявом пне березы (3 м). В них 3.07 находились выводки, которым оставалось 3–5 дней до вылета, в одном – 13 птенцов, в другом – 6 живых и 1 погибший. Судя по их возрасту, откладка яиц в гнездах началась в 5–6-й пятидневках мая (табл. 6). В 1976 г. первая птица встречена уже 5.05, начало массового пения и пара отмечены 12.05. Искусственные гнездовья заняли 3 пары, каждая из которых разорила 5–6 ближайших гнезд других дуплогнездников, отложив в 5 освобожденных синичников по 1–4 яйца. В одной из них тут же поселились большие синицы, построившие свое гнездо поверх яиц вертишеек. Первое яйцо появилось 29.05; кладки, которые птицы сели насиживать, начаты 3.06 (11 яиц), 6.06 (10 яиц) и 26.06 (7 яиц). Последняя, вероятно, была повторной: ранее вертишейки гнездились в соседнем гнездовье, куда к 16.06 отложили 3 яйца, вытащив из него гнездо большой синицы с птенцами. Однако к 20.06 эти яйца исчезли. Из гнезд с крупными кладками вылетело по 4 птенца. В первом из них одно яйцо не было оплодотворено, 4 птенца исчезли в период вылупления, еще 2 – после него, во втором – 5 яиц оказались без эмбрионов и один птенец исчез в дни вылупления. Основной причиной отхода птенцов могла быть большая разница в их возрасте, достигавшая 4–5 дней. Кроме того, судя по наличию во всех синичниках раковин, вертишейки кормили потомство довольно крупными наземными моллюсками, которые могли быть не слишком подходящей пищей для 1–2-дневных птенцов. История третьей кладки до конца не прослежена. В 1977 г. численность птиц упала. Факт их пребывания подтвердила только находка 3.07 четырех холодных яиц вертишеек на пустом дне дуплянке, в которой 15.06 еще находилось гнездо мухоловки-пеструшки с 6 холодными яйцами, вероятно, брошенными из-за беспокойства.
Обсуждение
Расхождение видов по экологическим нишам происходит по многим компонентам – трофическим, пространственным, временным и пр. (Одум, 1986). При этом в первую очередь идет разделение по местообитаниям, затем по пище и в меньшей степени по времени (Schoener, 1989, цит. по: Хлебосолов, 2002). Виды, находившиеся под наблюдением, занимают разные ниши, перекрывающиеся полностью по одному, но жизненно важному ресурсу – местам для гнездования, отсутствие которых делает репродукцию невозможной.
Уровень напряженности конкурентных отношений между вторичными дуплогнездниками во многом зависит от соотношения числа дупел и количества претендующих на них особей (Miller, 2010). В старовозрастных широколиственных лесах, несмотря на сравнительно высокую численность птиц, мест для их гнездования вполне достаточно благодаря обилию фаутного древостоя (Wesolowski, 2003, 2007). В приполярных лесах, напротив, и дупел, и птиц немного, поэтому при развеске дуплянок их численность увеличивается медленно и незначительно (Зацаринный, Константинов, 2007). В лесах, произрастающих на широтах южной Карелии, такого соответствия нет, и при привлечении дуплогнездников в искусственные гнездовья плотность их населения может возрастать многократно (Зимин, 1973; Артемьев, 2008 и др.). На о-ве Клименецком у большой синицы она увеличилась почти в 20, у мухоловки-пеструшки – в 15 раз.
Пик конкурентных отношений дуплогнездников приходится на начало гнездового периода, когда они распределяются по участкам и строят гнезда (Slagsvold, 1975; Krist, 2004). Пытаясь потеснить предыдущих хозяев, птицы устраивают драки, вытаскивают из дупел их гнезда или строят свои поверх яиц, отложенных конкурентами. Наблюдения показывают, что в регионах, где интервал между сроками гнездования конкурирующих видов достаточно велик, борьба идет в менее жестких формах. Там, где размножение проходит более дружно, она, как правило, ужесточается. Однако большие различия условий в разных местностях не позволяют при сравнении полностью исключить возможное влияние и других факторов, в том числе широтных и локальных особенностей фенологии (Slagsvold, 1976) или разную конструкцию искусственных гнездовий (Lambrechts et al., 2010; Møller et al., 2014). Использование данных, полученных в одной точке в смежные годы, различающиеся по характеру погодных условий, дает возможность сократить число подобных факторов.
Среди птиц, заселявших искусственные гнездовья на о-ве Клименецком, наибольшей агрессивностью отличаются большие синицы, охраняющие не только жилые гнезда, но и ближайшие пустые дуплянки, которые они используют для второго гнездования и ночлега самцов. В некоторых регионах во время драк они часто убивают мухоловок-пеструшек и других птиц (Slagsvold, 1975; Смирнов, 2012), но в Карелии такие случаи не известны. Вертишейки тоже способны убивать конкурентов, однако основной ущерб они наносят тем, что вытаскивают из соседних дупел их гнезда, иногда вместе с кладками и даже птенцами (Haartman, 1957; Головань, 1976 и др.). В ожесточенные, но обычно менее успешные схватки вовлекаются также мухоловки пеструшки и горихвостки (Tompa, 1967; Meidell, 1961; Merilä, Wiggins, 1995). Наличие защищаемых «резервных» дупел – одна из причин того, что на о-ве Клименецком, как и повсеместно, лишь половина синичников была занята жилыми гнездами, несмотря на избыток пустых гнездовий, наиболее привлекательные из них длительное время оставались объектами соперничества.
Сроки и ход размножения птиц зависят, прежде всего, от температурных и погодных условий региона, межгодовые колебания которых отражаются на всех фенологических явлениях. В южной Карелии у 62° с. ш. продолжительность безморозного периода варьирует в пределах трех месяцев (169–258 дней), сокращаясь в отдельные годы до показателей, обычных для Заполярья (Скороходова, 2008). Столь широкие колебания сказываются на устойчивости северных популяций птиц, поскольку здесь даже незначительное сокращение короткого благоприятного периода существенно ухудшает возможности их воспроизводства.
Для птиц особенно важны условия предгнездового периода. При раннем наступлении тепла представители видов, зимующих или рано прилетающих в Карелию, начинают кладки на 2–4 недели раньше, чем при позднем, что почти на треть удлиняет период репродукции их местных популяций. При наиболее низких температурах начинают гнездование большие синицы (Veistola et al., 1995). В южной Карелии даты появления их первых кладок варьируют в пределах месяца (Зимин, 1978; Artemyev, 2008).
Таблица 6. Сроки прилета и начала размножения дуплогнездников на о-ве Клименецкий в 1975–1976 гг.
Table 6. The time of arrival and breeding start of secondary hole-nesting birds on the island Klimenetsky, 1975–1976
| Вид | Дата первой встречи | Дата откладки первого яйца | ||
| 1975 | 1976 | 1975 | 1976 | |
| P. major | 30.04 | 10.05 | ||
| F. hypoleuca | 5.05 | 5.05 | 15.05 | 23.05 |
| Ph. phoenicurus | ? | 7.05 | 18.05 | 20.05 |
| J. torquilla | 8.05 | 5.05 | 20–23.05 | 29.05 |
Остальные виды – дальние мигранты, достигающие зимой Северной и Центральной Африки, перелеты которых регулируются в основном эндогенно (Дольник, 1975; Gwinner, 1996). В гнездовой области они появляются в относительно поздние и постоянные сроки без жесткой зависимости от местных условий (табл. 6). Сроки начала их гнездования колеблются не так сильно, что приводит к ежегодным изменениям интервала между началом размножения зимующих и мигрирующих видов, влияющего на напряженность межвидовых отношений (Ahola et al., 2007).
Экстремально теплой весной 1975 г. птицы приступили к размножению очень рано (Хохлова, 1976). При этом зимовавшие в регионе большие синицы успели отложить первые яйца до появления других дуплогнездников, а мухоловки-пеструшки начали кладки без обычной задержки, поскольку необходимый температурный порог был перейден до их прилета (рис. 1). В ранние сроки загнездились горихвостки и вертишейки (табл. 6). Холодной весной 1976 г. синицы начали откладку яиц на декаду позже и менее дружно. Пеструшки, появившиеся в обычные сроки, строили гнезда дольше и приступили к размножению на неделю позднее (Хохлова, Головань, 1981). Сдвинулся и пик их массовой кладки. Задержалось также начало репродукции основного разорителя гнезд мелких дуплогнездников – вертишейки.
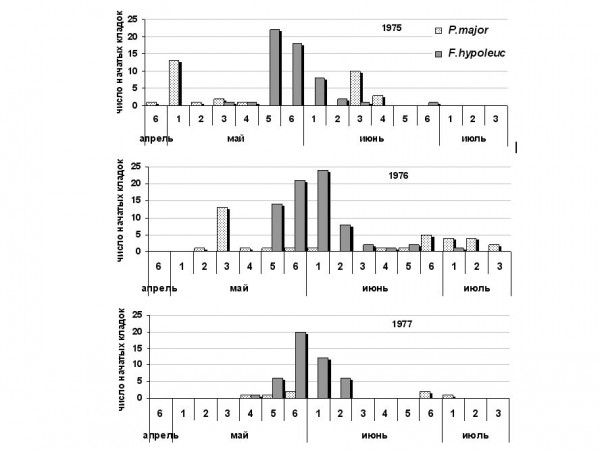
Рис. 1. Динамика размножения большой синицы P. major и мухоловки-пеструшки F. hypoleuca в 1975–1977 гг.
Fig. 1. The dynamics of reproduction of Great Tit P. major and Pied Flycatcher F. hypoleuca, 1975–1977.
Колебания сроков начала гнездования не повлияли на величину кладки и частоту вторых циклов большой синицы, которая имеет возможность начинать репродукцию в одной и той же фенологической ситуации. В отличие от синиц, мухоловка-пеструшка, попав в 1975 г. на более позднюю фенофазу, откликнулась резким увеличением плодовитости. Это же явление наблюдали в Финляндии, связав с необычайно ранним и массовым выплодом гусениц пяденицы Operopthera fagata (Pasanen, 1977). Но уже в следующем, 1976 г. сроки и величина кладки вернулись к обычным значениям.
Более значимо межгодовые колебания условий предгнездового периода отразились на успешности гнездования, что было связано в основном с вызванными ими изменениями структуры межвидовых отношений. В 1975 г. можно было наблюдать достаточно далекое расхождение экологических ниш разных видов дуплогнездников по времени. В середине мая, когда мигрирующие птицы распределялись по участкам и осматривали дуплянки, синицы уже сидели на яйцах, контролируя лишь ближайшие гнездовья, выбранные для второго цикла. Это защитило их кладки от разорения (погибло 2), а других птиц – от лишнего беспокойства. Возможно, по этой причине две пары вертишеек, поселившихся на участках синиц, заняли естественные дупла в стороне от дуплянок. В 1976 г. сроки размножения всех видов сблизились. Большие синицы приступили к репродукции уже после прилета мигрантов, в период, когда те интенсивно посещали дуплянки, устраивая драки и мешая друг другу. Особенно активно вели себя вертишейки. Из-за гибели гнезд 7 из 16 пар синиц вынуждены были трижды начинать кладки. Ситуацию усугубила и повысившаяся после успешного года численность всех дуплогнездников. О вероятном дефиците мест для размещения гнезд можно судить и по неоднократным случаям гнездования в одной дуплянке тех же пар или птиц других видов после вылета предыдущих выводков.
Повышенная напряженность конкурентных отношений между дуплогнездниками в год с поздней весной внесла весомый вклад в снижение успешности их репродукции. Усиление беспокойства вызвало рост числа гнезд, брошенных или разоренных во время постройки и откладки яиц, привело к появлению большого числа возобновленных кладок, меньших по величине и более поздних по срокам (табл. 3), вызвало дополнительный сдвиг средних сроков размножения. Все это способствовало падению продуктивности размножения популяций и увеличению необходимых репродуктивных усилий у значительной части конкурирующих особей.
Колебания численности птиц в 1975–1977 гг. соответствовали результатам гнездования в предыдущем году. У всех видов после успешного 1975 г. она выросла или, по крайней мере, не сократилась (большая синица), после менее удачного 1976 г. – снизилась (табл. 2). Известно, что птицы не всегда охотно заселяют искусственные гнездовья в год их развески. Не исключено, что этот фактор повлиял на показатели 1975 г., но эффект, по-видимому, не был значимым, поскольку те же тенденции в динамике численности этих видов отмечены на стационарах в Прионежье (Шокшинский) и восточном Приладожье (Гумбарицы), где работы проводили с 1970 г. (Зимин, Кузьмин, 1980; Хохлова, Головань, 1981). Особенно резкое сокращение численности (в 3 раза), которое невозможно объяснить одним уменьшением продуктивности гнездования, продемонстрировала большая синица. Причиной столь глубокого падения могло стать увеличение осенней смертности птиц из-за поздней смены оперения у значительной части популяции.
В южной Карелии постювенальная линька больших синиц начинается в возрасте 40–65 дней и продолжается не менее 50 дней (Рымкевич, Артемьев, 1990). Птенцы из поздних гнезд приступают к ней только в конце августа – начале сентября, отрастание нового оперения завершается в октябре – ноябре. В 1976 г. сложилась ситуация, крайне неблагоприятная для вида. Позднее наступление тепла задержало начало гнездования, сократив период возможной откладки яиц и восстановления разоренных гнезд, число которых выросло вследствие усиления конкуренции. И если в предыдущем году все кладки были начаты до 20.06, то в 1976 г. в половине гнезд яйца появились только в конце июня – середине июля. Это привело к увеличению доли поздних выводков, вынужденных переходить к самостоятельной жизни и менять оперение в сроки и условиях, далеких от оптимальных. Положение ухудшило раннее наступление холодов (Скороходова, 2006). Первый заморозок у 62° с. ш. отмечен уже 30 августа, снегопад с установлением снежного покрова – 19.09, переход среднесуточных температур через 0 °С – в начале октября, когда у многих особей старое перо уже выпало, а новое еще полностью не отросло.
Смена наряда требует больших энергетических затрат. Кроме того, во время линьки птицы очень чувствительны к качественному составу кормов (Дольник, 1975). Ее сдвиг на период быстрого ухудшения погодных и кормовых условий отражается на уровне смертности, что на севере может делать вторые выводки больших синиц невыгодными (Orell, Ojanen, 1986). В Приладожье при высокой осенней численности большой синицы (более 46.2 ос./км²), характерной и для о. Клименецкого, плотность гнездового населения прямо коррелирует со среднемесячной температурой ноября и мало зависит от зимних температур (Artemyev, 2008). Это свидетельствует о том, что поздние сроки линьки, закрепленные генетически, могут служить серьезным препятствием для устойчивого существования северных популяций видов с полицикличным размножением. Подтверждением служат и результаты 30-летних наблюдений за экспансией черного дрозда T. merula L. на северо-восток, успешное закрепление которого в регионе сопровождалось постепенным смещением линьки на более ранние сроки (Khokhlova, 2009). Позже других заканчивают смену оперения самки больших синиц, выкормившие вторые и поздние повторные выводки: не завершившие ее особи встречаются даже в начале декабря (Рымкевич, Артемьев, 1990). Высказывается предположение, что на севере наличие двух циклов размножения у этого вида «не только не способствует приросту популяции за счет большого количества молодых, но и ставит в неблагоприятные условия взрослых птиц, имеющих две кладки» (Шутова, 2001, стр. 659).
Таким образом, резкий межгодовой перепад весенних температур в 1975–1976 гг. повлиял на многие популяционные характеристики вторичных дуплогнездников не только напрямую, но и опосредованно, через изменение напряженности межвидовых отношений. Усиление конкуренции в год с холодной весной сыграло заметную роль в снижении успешности гнездования и увеличении смертности птиц в послегнездовой период.
Заключение
В северных широтах сроки прилета и размножения птиц повсеместно варьируют по годам под влиянием изменений природной обстановки. Согласно теории экологических ниш, это не может не отражаться на межвидовых отношениях птиц, использующих один ресурс. Однако эти колебания обычно не заметны из-за небольшой величины межгодовых сдвигов. Анализ данных, полученных в Карелии в смежные годы с экстремально разным характером весны, позволил подтвердить, что в нестабильных условиях севера конкуренция может вносить весомый вклад в динамику популяционных характеристик птиц и выступать как один из значимых элементов механизма регуляции их численности. Изменения ее напряженности при резком межгодовом перепаде весенних температур значимо отразились на продуктивности размножения и смертности вторичных дуплогнездников, что способствовало соответствующим изменениям гнездовой плотности в последующие годы.
Библиография
Артемьев А. В. Популяционная экология мухоловки-пеструшки в северной зоне ареала . М.: Наука, 2008. 267 с.
Головань В. И. О географических особенностях размножения мухоловки–пеструшки // Материалы 9–й Прибалт. орнитол. конф. Вильнюс, 1976. С. 72–75.
Дольник В. Р. Миграционное состояние птиц . М.: Наука, 1975. 399 с.
Зацаринный И. В., Константинов В. М. Особенности размножения птиц-дуплогнездников на северной границе распространения сосновых лесов // Рус. орнитол. журн. 2007. Экспресс-вып. Т. 16. № 353. С. 471–485.
Зимин В. Б. Итоги работ по привлечению птиц-дуплогнездников // Тр. гос. заповедника «Кивач». Петрозаводск, 1973. Вып. 2. С. 40–63.
Зимин В. Б. Материалы по гнездованию большой синицы (P. major L.) в Карелии // Фауна и экология птиц и млекопитающих таежного Северо–Запада СССР. Петрозаводск, 1978. С. 17–31.
Зимин В. Б., Ивантер Э. В. Фаунистический обзор наземных позвоночных заповедника «Кивач» // Труды заповедника «Кивач». Вып. 1. Петрозаводск, 1969. С. 22–64.
Зимин В. Б., Кузьмин И. А. Экологические последствия применения гербицидов в лесном хозяйстве . Л.: Наука, 1980. 175 с.
Зимин В. Б., Сазонов С. В., Лапшин Н. В., Хохлова Т. Ю., Артемьев А. В., Анненков В. Г., Яковлева М. В. Орнитофауна Карелии . Петрозаводск: Изд. КНЦ РАН, 1993. 220 с.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск: Изд-во ПетрГУ, 2003. 304 с.
Лакин Г. Ф. Биометрия . М.: Высшая школа, 1990. 352 с.
Одум Ю. Экология . М.: Мир, 1986. Т. 2. 376 с.
Пианка Э. Эволюционная экология . М.: Мир, 1981. 400 с.
Розенберг Г. С., Рянский Ф. Н. Теоретическая и прикладная экология . Нижневартовск: Изд-во Нижневарт. пед. ин-та, 2005. 292 с.
Рымкевич Т. А., Артемьев А. В. Большая синица (Parus major) // Линька воробьиных птиц Северо–Запада СССР. Л.: Изд-во ЛГУ, 1990. С. 169–177.
Скороходова С. Б. Календарь природы заповедника «Кивач» (1966–2005 гг.) // Труды зап. «Кивач». Вып. 3. Петрозаводск, 2006. С. 48–79.
Скороходова С. Б. О климате заповедника «Кивач» // Труды зап. «Кивач». Вып. 4. Петрозаводск, 2008. С. 3–34.
Смирнов О. П. О необычных взаимоотношениях большой синицы Parus major с некоторыми видами птиц-дуплогнездников в сезон размножения // Рус. орнитол. журн. 2012. Экспресс-вып. Т. 21. № 732. С. 435–438.
Хлебосолов Е. И. Экологические факторы видообразования у птиц . М.: Горизонт, 1999. 284 с.
Хлебосолов Е. И. Теория экологической ниши: история и современное состояние // Русский орнитологический журнал. 2002. Экспресс-вып. Т. 11. № 203. С. 1019–1037.
Хохлова Т. Ю. Фенология прилета и размножения птиц в условиях ранней весны // Экология птиц и млекопитающих северо-запада СССР. Петрозаводск: Изд-во КФАН СССР, 1976. С. 38–44.
Хохлова Т. Ю. Горихвостка-лысушка Phoenicurus phoenicurus (L.) // Красная книга Карелии / Ред Э. В. Ивантер, О. Л. Кузнецов. Петрозаводск: Карелия, 1995. С. 180–181.
Хохлова Т. Ю., Головань В. И. К биологии размножения мухоловки-пеструшки в Южой Карелии // Экология наземных позвоночных Северо–Запада СССР / Под ред. Э. В. Ивантера. Петрозаводск: Изд-во КФАН СССР, 1981. С. 50–62.
Шутова Е. В. О вторых кладках большой синицы на севере европейской части России // Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии: Материалы Междунар. конф. (ХI Орнитологическая конф.). Казань: Матбугат йорты, 2001. С. 658–659.
Ahola M., Laaksonen T., Eeva T., Lehikoinen E. Climate change can alter competitive relationships between resident and migratory hole–breeders // Journal of Animal Ecology. 2007. Vol. 76. P. 1045–1052.
Artemyev A. V. Population ecology of the Great Tit Parus major in the taiga forest on Lake Ladoga coast // Avian Ecology and Behaviour. 2008. Vol. 14. P. 1–33.
Gause G. F. The Struggle for Existence. Baltimore: Williams and Wilkins, 1934. 163 p.
Gwinner E. Circannual clocks in avian reproduction and migration // Ibis. 1996. Vol. 138. P. 47–63.
Haartman L. von. Adaptation in hole-nesting birds // Evolution. 1957. Vol. 11. P. 339–347.
Haartman L. von. Population dynamics // Farner D. S., King J. R. (Eds.). Avian Biology. London: Academic Press, 1971. Vol. 1. P. 391–459.
Khokhlova T. Y. Juvenile moult and spatial behaviour of first–year Blackbirds Turdus merula on the northeast edge of the range // Avian Ecology and Behaviour. 2009. Vol. 15. P. 1–22.
Krist M. Importance of competition for food and nest–sites in aggressive behaviour of Collared Flycatcher Ficedula albicollis // Bird Study. 2004. № 51. P. 41–47.
Lambrechts M. M., Adriaensen F., Ardia D. R., Artemyev A., Attenzar F., Banbura J., Barba E. et al. The design of artificial nest boxes for the study of secondary hole–nesting birds: a review of methodological inconsistencies and potential biases // Acta Ornithol. 2010. Vol. 45. P. 1–26.
Meidell O. Life history of the Pied Flycatcher and the Redstart in a Norwegian mountain area // Nytt Mag. Zool. 1961. Vol. 10. P. 5–48.
Merilä J., Wiggins. D. A. Interspecific competition for nest holes causes adult mortality in the Collared Flycatcher // The Condor. 1995. Vol. 97. P. 445–50.
Miller K. E. Nest-Site Limitation of Secondary Cavity-Nesting Birds in Even-Age Southern Pine Forests // The Wilson Journal of Ornithology. 2010. Vol. 122. № 1. P. 126–134.
Minot E. O., Perrins C. M. Interspecific competition–nest sites for Blue and Great Tits // J. Anim. Ecol. 1986. Vol. 55. P. 331–350.
Møller A. P., Adriaensen F., Artemyev A., Banbura J., Biard C. et al. Clutch size in European secondary hole–nesting passerines in relation to nest–box floor area, habitat, geographic location and study year // Methods Ecol. Evol. 2014. Vol. 5. P. 353–362.
Newton I. The role of nest sites in limiting the numbers of hole–nesting birds: A review // Biol. Conserv. 1994. Vol. 70. P. 265–276.
Newton I. Population Limitation in Birds. London: Academic Press, 1998. P. 167–187.
Orell M., Ojanen M. Relation of fitness of female Great Tits to clutch size and number of broods // Ornis Fenn. 1986. Vol. 63. P. 120–127.
Pasanen S. Breeding biology of the pied flycatcher (Ficedula hypoleuca) in eastern Finland // Ornis Fennica. 1977. Vol. 54. № 3. P. 119–122.
Slagsvold T. Competition between the great tit Parus major and the pied flycatcher Ficedula hypoleuca in the breeding season // Ornis Scandinavica. 1975. Vol. 6. P. 179–190.
Slagsvold T. Annual and geographical variation in the time of breeding in the Great Tit Parus major and the Pied Flycatcher Ficedula hypoleuca in relation to environmental phenology and spring temperature // Ornis Scandinavica. 1976. Vol. 7. P. 127–145.
Slagsvold T. Competition between the Great Tit Parus major and the Pied Flycatcher Ficedula hypoleuca: An Experiment // Ornis Scandinavica. 1978. Vol. 9. № 1. P. 46–50.
Tompa F. S. Reproductive success in relation to breeding density in the Pied Flycatchers, Ficedula hypoleuca (Pallas) // Acta zool. Fenn. 1967. Vol. 118. P. 1–28.
Vallin N., Rice A. M., Arntsen H., Kulma K., Qvarnström A. Combined effects of interspecific competition and hybridization impede local coexistence of Ficedula flycatchers // Evolutionary Ecology. 2012. Vol. 26. P. 927–942.
Veistola S., Lehikoinen E., Iso-Iivari L. Breeding biology of the Great Tit Parus major in a marginal population in Northernmost Finland // Ardea. 1995. Vol. 83. P. 419–420.
Wesołowski T. Bird community dynamics in a primaeval forest – is interspecific competition important? // Ornis Hung. 2003. Vol. 12–13. P. 51–62.
Wesołowski T. Lessons from long–term hole–nester studies in a primeval temperate forest // J. Ornithol. 2007. Vol. 148 (Suppl 2). P. 395–405.
Благодарности
Благодарю Н. В. Морошенко, Н. Л. Кольцову, Л. В. Коротецкову, Л. В. Фирсову, Ф. А. Карлика, В. Г. Высоцкого и многих других коллег, студентов и юннатов, принимавших участие в полевых работах в Кижских шхерах Онежского озера в 1970-е годы. Выражаю особую признательность М. В. Яковлевой, помогавшей на всех этапах работы, и А. В. Артемьеву за участие в обсуждении и редактировании статьи.

 © 2011 - 2024
© 2011 - 2024