



Введение
У большинства особей хозяина в течение жизни встречается более одного вида паразитов. Эти паразитарные сообщества (инфрасообщества) характеризуются высокой изменчивостью и формируются под действие внутренних (например, иммунный, гормональный, физиологический статус хозяина) и внешних (сезонность, тип местообитания, особенности поведения хозяина и др.) факторов (Cattadori et al., 2006). Помимо этого, на структуру инфрасообществ могут оказывать влияние и межвидовые взаимодействия между паразитами.
Можно предположить, что адаптация к совместному паразитированию, главным образом, должна сводиться к уравновешиванию патогенного действия комплекса паразитов с иммуно-физиологическими возможностями хозяина. В связи с этим, при анализе паразитарного сообщества любого хозяина могут наблюдаться отрицательные зависимости между разными видами паразитов. К примеру, возрастание численности одного вида, в соответствие с принципом конкурентного исключения, приведет к снижению другого.
Целью нашего исследования был анализ межвидовых взаимоотношений паразитов рыжей полевки, направленный на выявление связей между показателями зараженности паразитами разных экологических и систематических групп.
Материалы
Материалом для исследований послужили сборы паразитов мышевидных грызунов, выполненные в мае-октябре 1994– 2000 гг. в районе стационара ИБ КарНЦ РАН (62º04′ с.ш, 33º55′ в.д) в среднетаежной подзоне Карелии. Мелких млекопитающих отлавливали линями ловушек Геро. При сборе и обработке материала применяли стандартные зоологические и паразитологические методики (Жмаева и др., 1964; Коросов, 1994; Аниканова и др., 2010).
По степени развития генеративной системы выделяли 4 основных класса зрелости мышевидных грызунов (Коросов, 1994): молодые, взрослые, созревающие, старые. Молодые (неполовозрелые) самки имеют нитевидную матку, влагалище – без пробки; у самцов – мелкие семенники (3 мм) и придатки (1 мм). У взрослых (половозрелых) самок матка утолщена, во влагалище после оплодотворения появляется хрящевая пробка, на поздних стадиях беременности видны эмбрионы, у рожавших на рогах матки остаются плацентарные пятна. У самцов увеличиваются семенники (11–12 мм). Созревающие животные имеют промежуточные размеры половых органов, к размножению не способны. Старые особи имеют дегенерировавшие половые органы темного цвета, по размерам большие, чем у молодых; к размножению не способны. Зрелость животного рассматривали как критерий возраста.
Всего проанализированы данные по 409 экз. рыжей полевки Myodes glareolus (Schreber, 1780), из которых 217 молодых, 25 созревающих и 167 взрослых животных. Возраст молодых животных составил 1–2 месяца, взрослых – 6–10 мес. (зимовавшие полевки) и 3–4 мес. (сеголетки первой генерации).
При паразитологическом вскрытии животного подсчитывали число (шт.) особей каждого вида эндо- и эктопаразитов. Отдельная особь рыжей полевки – это единица местообитания N-го количества видов паразитов с n-ой численностью.
Методы
Соответствие эмпирических данных значений паразитарной нагрузки (количество всех паразитов на одной особи хозяина) с основными законами распределения оценивали по критерию χ2.
Для оценки конкурентной напряженности в сообществе паразитов рыжей полевки применяли индекс C-score (Stone, Roberts, 1990). Показатель C-score рассчитывают как среднее значение числа "единиц шахматной доски" для всех пар видов в наборе данных. Число единиц шахматной доски (CU) для каждой пары видов вычисляют как
CU = (ri – S) (rj – S),
где S – число общих мест (особь хозяина, где встречаются оба вида), ri и rj – количество рядов (животных) с паразитом i и j. В нашем случае единица шахматной доски имеет вид:
Полевка 1 Полевка 2
Паразит 1 0 1
Паразит 2 1 0
где 1 – вид отмечен; 0 – вид отсутствует.
Значение полученного индекса C-score сравнивают с ожидаемым, который рассчитывается при условии, что виды в сообществе распределены случайно независимо друг от друга. Если эмпирический C-score больше теоретического, то наблюдается сегрегация видов (некоторые виды встречаются менее часто, чем ожидается), что может быть следствием их конкуренции. И наоборот, если эмпирический C-score меньше теоретического, в сообществе наблюдается определенная агрегированность некоторых видов, что в свою очередь, может быть связано с их сходной реакцией на действие какого-либо фактора. Расчеты проводили в программе EcoSim (Gotelli, Entsminger, 2004).
Для оценки зависимости численности паразитов в смешанных инвазиях попарно рассчитывали ранговый коэффициент корреляции Спирмена (Ивантер, Коросов, 1992) массовых видов: нематод – Heligmosomum mixtum, Heligmosomoides glareoli, Syphacia petrusewiczi; иксодовых клещей – Ixodes trianguliceps, I. persulcatus; гамазовых клещей – Echinonyssus isabellinus, Haemogamasus nidi, Eulaelaps stabularis; блох – Ctenophthalmus uncinatus, Megabothris rectangulatus, Peromyscopsylla silvatica; у трех групп полевок.
Расчеты значений индексов C-score и r Спирмена проводили для трех групп полевок: случайная выборка, половозрелые животные и зрелые самцы.
Результаты
Анализ распределения значений паразитарной нагрузки в популяции рыжей полевки показал, что наблюдаемые эмпирические частоты логарифма численности совокупности паразитов согласуются с законом логнормального распределения χ2 = 11.1, df = 7, p = 0.14 (рис. 1). Характер кривой определяется наличием в популяции хозяина особей с высокой зараженностью, что может быть и результатом взаимодействия разных видов паразитов.
Значения наблюдаемого индекса C-score для трех групп рыжей полевки (случайная выборка, половозрелые особи и половозрелые самцы) значимо не отличались от теоретических, рассчитанных при допущении, что паразиты распределены случайным образом независимо друг от друга (табл. 1). Следовательно, при анализе встречаемости видов нами не выявлено ни конкурентного, ни синергетического действия паразитов друг на друга.
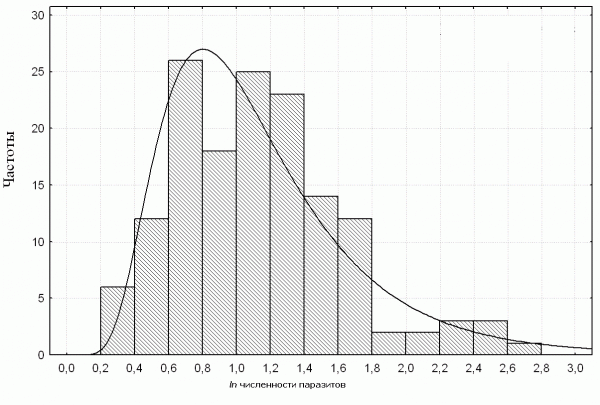
Рис. 1. Согласование частот распределения логарифма общей численности паразитов у рыжей полёвки с логнормальным типом распределения. Гистограмма – эмпирические; график – теоретические частоты
Fig. 1. The observed frequency distribution (histogram) and the probability lognormal distribution (curve) of parasite burdens in the bank vole.
Таблица 1. Расчеты индекса C-score для разных групп рыжей полевки Myodes glareolus
| Случайная выборка | Половозрелые особи | Половозрелые самцы | |
| N рыжей полевки | 255 | 167 | 86 |
| Значение наблюдаемого индекса | 177.3 | 101.5 | 43.5 |
| Среднее значение ожидаемого индекса (при условии, что виды распределены независимо друг от друга) | 177.8 | 101.3 | 43.5 |
| Дисперсия ожидаемого индекса | 1.24 | 0.62 | 0.18 |
| p (наблюдаемый ≤ ожидаемого) | 0.34 | 0.54 | 0.48 |
| p (наблюдаемый ≥ ожидаемого) | 0.66 | 0.46 | 0.52 |
В выборке, составленной из всех животных, значимые коэффициенты ранговой корреляции Спирмена (р < 0.05) получены для 10 пар паразитов из 55 проанализированных (табл. 2). При разделении животных по полу и возрасту наблюдается снижение числа корреляций. Так, при анализе выборки из половозрелых особей и зрелых самцов значимые зависимости получены для 7 пар паразитов в каждой группе (табл. 3, 4). Значимые отрицательные корреляции не выявлены, все наблюдаемые зависимости были положительными. В трех выборках общими парами паразитов, для которых отмечена положительная корреляция, были: Ixodes persulcatus – I. trianguliceps (иксодовые клещи), Haemogamasus nidi – Eulaelaps stabularis (гамазовые клещи) и Eulaelaps stabularis – Megabothris rectangulatus, Peromyscopsylla silvatica (блохи). Если у некоторых взрослых (более старших) особей указанные пары паразитов могли одновременно встречаться, то у молодых они одновременно, как правило, отсутствовали. Это обстоятельство и породило положительную корреляцию в смешанной группе животных.
Таблица 2. Коэффициент ранговой корреляции Спирмена численности паразитов рыжей полевки
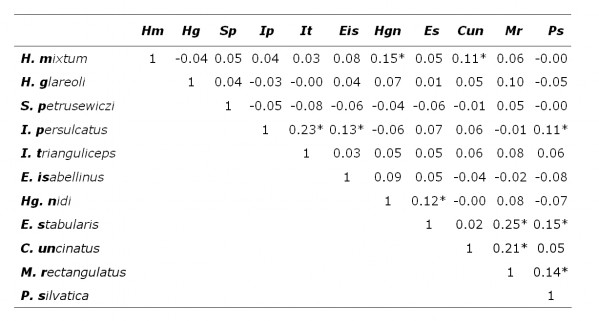
* – значимо отличается от нуля при р < 0.05
Таблица 3. Коэффициент ранговой корреляции Спирмена численности паразитов рыжей полевки (половозрелые особи)
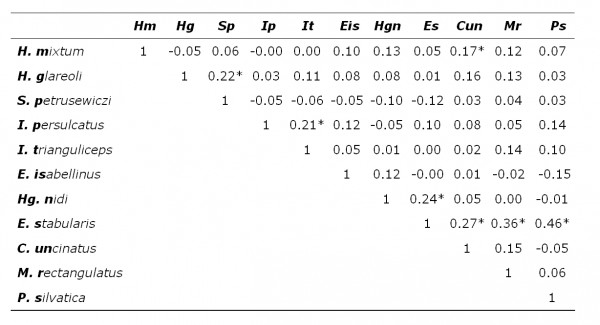
* – значимо отличается от нуля при р < 0.05
Таблица 4. Коэффициент ранговой корреляции Спирмена численности паразитов рыжей полевки (половозрелые самцы)
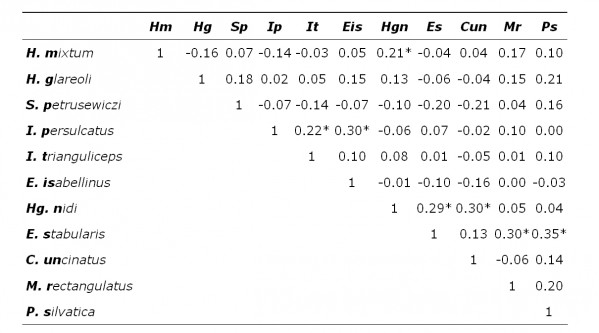
* – значимо отличается от нуля при р < 0.05
Обсуждение
Согласование полученных данных по индивидуальной зараженности рыжей полевки с логнормальным типом распределения может быть связано с асиметричным действием некоторых факторов, приводящих к возникновению характерного «хвоста» гистограммы. Наблюдаемая правосторонняя асимметрия обусловлена наличием в популяции хозяина гиперинвазированных особей, и может быть результатом определенного синергизма паразитов разных видов, когда высокая зараженность каким-либо видом приводит к снижению иммунитета хозяина, и делает его более доступным для других паразитов.
Выполненный нами анализ совместной встречаемости (индекс C-score) не показал ни синергетического, ни конкурентного действия паразитов в смешанных инвазиях. Присутствие любого вида в хозяине не сказывается на вероятности обнаружения другого (табл. 1). Однако при анализе количественных данных по численности массовых видов паразитов получено большое число значимых положительных коэффициентов корреляции, которые в свою очередь и отражают наблюдаемую асимметрию распределения паразитов (рис. 1). Отмеченные нами корреляционные связи, видимо, обусловлены сходной реакцией (например, рост или снижение численности) разных видов на действие какого-либо фактора.
К таким факторам, очевидно, относится пол и возраст хозяина. Анализ паразитофауны разных половозрастных групп рыжей полевки на территорию южной Карелии показал, с одной стороны, увеличение зараженности с возрастом животных, с другой – большую зараженность половозрелых самцов по сравнению с самками (Бугмырин, 2003). Следовательно, наблюдаемые положительные корреляции могли возникать вследствие того, что у старших особей указанные пары паразитов одновременно встречались, а у молодых они, как правило, отсутствовали. Для нивелирования влияния пола и возраста хозяина на совместную встречаемость паразитов мы рассчитали коэффициенты корреляций отдельно для зрелых полевок (исключили возраст) и зрелых самцов (исключили пол и возраст). И в том и в другом случае появились новые положительные зависимости, а общее число корреляций уменьшилось. Соответственно, можно предположить, что те корреляции, которые наблюдались при анализе всей выборки животных и не проявились при расчетах у половозрастных групп, отражали как раз связь паразитов с этими факторами. Таким образом, общее снижение числа положительных корреляций может свидетельствовать и об отсутствии прямого синергетического действия паразитов разных видов друг на друга в смешанных инвазиях, когда все наблюдаемые положительные корреляция – лишь результат влияния каких-либо неучтенных внешних условий, однонаправленных для данных паразитов.
Другими авторами получены аналогичные результаты для гельминтов (Montgomery, Montgomery, 1990; Haukisalmi, Henttonen, 1993) и эктопаразитов (Lundqvist, Brinck-Lindroth, 1990) мышевидных грызунов. В этих работах говорится о преобладании нейтральных и положительных межвидовых отношений в паразитарных сообществах. Отрицательные ассоциации гельминтов рыжей полевки были показаны только у видов одной таксономической группы (нематоды или цестоды), имеющих одинаковую локализацию (Kisielewska, 1970). Возможно, отрицательные связи имеют место в более южных районах, где паразитофауна рыжей полевки характеризуется большим разнообразием.
Заключение
Наши исследования межвидовых взаимоотношений паразитов рыжей полевки не выявили антагонистического действия паразитов друг на друга. Наблюдаемые положительные зависимости могут определяться сходной, независимой друг от друга, реакцией разных видов паразитов на изменения среды I (организм хозяина) или II (место обитания хозяина) порядка.
Библиография
Аниканова В. С., Бугмырин С. В., Иешко Е. П. Методы сбора и изучения гельминтов мелких млекопитающих: Учебное пособие. Петрозаводск: КарНЦ РАН, 2007. 145 с.
Бугмырин С. В. Эколого-фаунистический анализ паразитов мышевидных грызунов южной Карелии. : Автореф. дис. … канд. биол. наук. Петрозаводск, 2003. 18 с.
Жмаева З. М., Земская А. А., Шлугер Е. Г. Кровососущие клещи (Arthropoda, Frachnoidea, Chelicerata): Общие вопросы сбора и обработки материалов. // Методы изучения природных очагов болезней и человека / Под ред. П. А. Петрищевой, Н. Г. Олсуфьева. М.: Медицина, 1964. С. 68–73.
Ивантер Э. В., Коросов А. В. Основы биометрии: Введение в статистический анализ биометрических явлений и процессов: Учебное пособие. Петрозаводск: ПетрГУ, 1992. 168 с.
Коросов А. В. Организация летней практики по зоологии позвоночных животных:Учебное пособие. ПГУ. Петрозаводск, 1994. 67 с.
Cattadori I. M., Haukisalmi V., Henttonen H., Hudson P.J. Transmission ecology and the structure of parasite communities in small mammals. In: Micro mammals and macroparasites: from evolutionary ecology to management (Morand S., Krasnov B, Poulin R. eds.).Springler-Verlag, 2006. P. 349–369.
Gotelli N. J., Entsminger G. L. EcoSim: Null models software for ecology. Version 7. 2004. Acquired Intelligence Inc. & Kesey-Bear. Jericho, VT 05465. URL: http://homepages.together.net/~gentsmin/ecosim.htm (дата обращения: 29.11.2002).
Haukisalmi V., Henttonen H. Co-existence in helminths of the bank vole Clethrionomys glareolus. I. Patterns of co-occurrence // Journal of Animal Ecology. Vol. 62. N 2. 1993. P. 221–229.
Kisielewska K. Ecological organization of intestinal helminth groupings in Clethrionomys glareolus (Schreb.) (Rodentia). V. Some questions concerning helminth groupings in the host individuals // Acta Parasitologica Polonica. Vol. 17. 1970. P. 197–208.
Lundqvist L., Brinck-Lindroth G. Patterns of coexistence: ectoparasites on small mammals in northern Fennoscandia // Ecography. Vol. 13. N 1. 1990. P. 39–49.
Montgomery S. S. J., Montgomery W.I., Structure, stability and species interactions in helminth communities of wood mice Apodemus sylvaticus // International Journal for Parasitology. Vol. 20. N 2. 1990. P. 225–242.
Schluter D. A variance test for detecting species associations, with some example applications // Ecology. Vol. 65. N 3. 1984. P. 998–1005.
Stone L., Roberts A. The checkerboard score and species distributions // Oecologia. Vol. 85. N 1. 1990. P. 74–79.
Благодарности
Авторы выражают признательность В. С. Аникановой за помощь в сборе и анализе паразитологического материала, Е. П. Иешко за общую организацию и руководство и А. В. Коросову за обсуждение результатов.
Исследования выполнены при поддержке Министерства образования и науки Российской Федерации, соглашение 8101.

 © 2011 - 2024
© 2011 - 2024