



Введение
К современному этапу развития науки о ландшафте в ней обозначились две магистральных линии исследований структуры ландшафта (Современное…, 2018). Первая линия концентрируется на выявлении факторов пространственной дифференциации, способах идентификации и картографирования пространственных единиц, изучении адаптации многообразных природных и социально-экономических объектов и явлений к пространственной структуре. Вторая линия сосредотачивает внимание на межкомпонентных радиальных связях, круговоротах, цепных реакциях между компонентами. Обе линии исследований могут содержать временную составляющую, описывающую динамику и эволюцию пространственных структур или набора свойств компонентов ландшафта. В последние годы своей деятельности Ю. Г. Пузаченко особенно активно развивал геодинамическое направление ландшафтоведения, изучающее механизмы пространственно-временной динамики и организации ландшафта и его компонентов (Пузаченко, 2014). Однако явным пробелом остается сюжет, который мог бы связать эти две линии между собой: зависимость динамики от характера пространственного контекста. Возможно, это связано с трудностью организации стационарных исследований, но в то же время новые возможности предоставляет доступность накопленной базы космических снимков и цифровых моделей рельефа.
Достигнутый уровень развития ландшафтоведения требует перехода от этапа идентификации, описания, классификации пространственных структур к этапу объяснения механизмов формирования эмерджентных свойств геосистем как результата взаимодействия элементов пространственной структуры. Решение этой задачи позволяет стабилизировать режимы функционирования природно-хозяйственных систем путем как подбора оптимальных площадных соотношений, конфигураций и соседств хозяйственный угодий, так и адаптации допустимых нагрузок к естественному диапазону изменчивости свойств ландшафта во времени.
Под стабильностью функционирования геосистемы в данной работе понимается стабильность принадлежности к тому или иному классу динамических изменений. В качестве индикатора функционирования мы используем нормализованный разностный вегетационный индекс (NDVI) как индикатор зеленой надземной фитомассы. В качестве меры динамических изменений рассматривается приращение (с положительным или отрицательным знаком) значения индекса между сроками наблюдений и мера отклонения приращения для фации от фонового приращения для ландшафта.
Изменчивость фации, по гипотезе, отражает не только ее собственные свойства, но состояние геосистемы более высокого ранга. По мнению Ю. Г. Пузаченко, взаимодействие обычно имеет довольно малую дальность с получением информации прежде всего от ближних соседей, что не устраняет всесторонне влияние поведения системы в целом (Пузаченко, 2017а). На изменчивость свойств может влиять пропорция, конфигурация, размеры, соседства пространственных элементов вышестоящей (более высокого ранга) геосистемы. С точки зрения ландшафтоведения и ландшафтной экологии этот аспект требует учета одновременного влияния разнообразных типов пространственных структур (Turner, Gardner, 2015; Гродзинський, 2015) и разделения их вкладов. В качестве вышестоящих, по отношению к фации, рассматриваются три типа ландшафтных структур (по В. Н. Солнцеву, 1997): геостационарные (обусловленные стабильными геолого-геоморфологическими характеристиками), геоциркуляционные (порождаемые гравитационными потоками вещества), биоциркуляционные (порождаемые почвенно-фитоценотическими отношениями в зависимости от тепло- и влагообеспеченности). При исследовании биоциркуляционных структур особенное значение приобретают положительные обратные связи между почвой и фитоценозом, которые позволяют преодолевать рамочные условия функционирования, заданные морфолитогенной основой.
Одной из важных граней исследований взаимодействия пространственной единицы с соседними и удаленными единицами считается оценка вклада позиционных факторов. Например, зависимость NDVI и его изменчивости от положения по отношению к соседним экологически значимым объектам недавно описана в гидролесомелиоративных исследованиях (Пахучий, Пахучая, 2014). В прикладном аспекте часто исследовалась изменчивость урожайности сельскохозяйственных культур (обычно по значениям NDVI) в зависимости от пространственной вариабельности рельефа и почв на полях, особенно в связи с разработкой технологий точного земледелия (Corwin, 2013; Kaspar et al., 2003; Basso et al., 2001; Verhulst, Govaerts, 2010).
Целью исследования было установить пространственные и позиционные условия существования разных классов стабильности функционирования. Предварительно необходимо было решить задачу о факторах пространственного варьирования надземной фитомассы как таковой и об изменчивости вкладов этих факторов во времени. Затем проверялись гипотезы о зависимости изменчивости фитомассы во времени от принадлежности к видам урочищ, положения фации внутри урочищ, соседств и конфигураций урочищ.
Материалы
Исследование проведено в низкогорно-степном ландшафте Южного Урала (левобережье р. Урал) на участке «Айтуарская степь» государственного заповедника «Оренбургский» и примыкающих территориях (рис. 1). Общая площадь исследования – 139 кв. км.

Рис. 1. Географическое положение территории исследования в Кувандыкском районе Оренбургской области (А) и ландшафтная структура заповедного участка (Б). 1–33 – номера видов урочищ (Хорошев, 2016)
Fig. 1. Geographical position of the study area in the Kuvandyk district, Orenburg region (А) and landscape pattern of the nature reserve (Б). 1–33 – stow types (Khoroshev, 2016)
Основная часть территории Айтуарской степи (западный и центральный сектора до западной окраины балки Тышкак) принадлежит ландшафту структурно-эрозионного складчатого грядово-балочного низкогорья, сложенного чередующимися пластами песчаников, конгломератов, известняков и алевролитов, с останцами эоценовых поверхностей выравнивания, с петрофитными разнотравно-ковыльно-типчаковыми степями на черноземах южных маломощных сильнокаменистых поверхностно-карбонатных (Хорошев, Леонова, 2015; Хорошев, 2016).
Ландшафтные описания производились на площадках 10 х 10 м в фациях репрезентативных для всех звеньев катен и секторов эрозионных форм с разной площадью водосбора. Общее количество описаний за период 2011–2019 гг. составило 216. В июне 2019 г. на 64 площадках 0.5 x 0.5 м были проведены укосы травяной надземной фитомассы. Укосы собирались в полиэтиленовые пакеты, после маршрута сразу же взвешивались в сыром состоянии; после высушивания взвешивались повторно для расчета сухой фитомассы и содержания влаги. Аналогичным образом определялась влажность гумусовых горизонтов почв, отобранных в бюксы.
Для решения задачи о пространственных факторах изменчивости зеленой надземной фитомассы во времени использованы данные NDVI, рассчитанные для 11 весенних и летних сроков съемки спутника Landsat 8 2014–2019 гг. с разрешением 30 м. Для характеристики рельефа использована цифровая модель SRTM (исходное разрешение 90 м) после процедуры даунскейлинга до разрешения 30 м методом Bicubic Spline Interpolation. Общее число пикселов составило 154703. Расчеты морфометрических показателей рельефа, классификация космического изображения производились средствами SAGA GIS 7.4.0.
Методы
Пространственное распределение растительных сообществ выявлялось путем классификации серии разносезонных многоканальных космических снимков Landsat 8 методом к-средних в программе SAGA GIS и полевой верификации результатов формализованной классификации. Разрешение снимка 30 м примерно соответствует размеру природно-территориального комплекса ранга фации.
Значения NDVI были рассчитаны по снимкам Landsat 8. Во избежание влияния климатических трендов использованы доступные снимки Landsat с датой съемки, отстоящей от полевых наблюдений не более чем на 5 лет, т. е. 2014–2019 гг. Для проверки гипотезы о зависимости условий биопродуцирования от рельефа средствами программы Statistica 7.0 построены мультирегрессионные линейные «геоморфологические» модели, в которых в качестве зависимой переменной выступали либо значения NDVI, либо характеристики их изменчивости во времени, а в качестве независимых – характеристики рельефа. Для этого в программе SAGA GIS рассчитаны: позиционный индекс TPI и индекс расчлененности TRI (для окрестностей 90, 150, 210, 270, 310, 390 м), уклон, экпозиция склона (выраженная показателями положения на осях «север – юг» и «запад – восток» через косинус и синус румба, измеренного в радианах соответственно), индекс конвергенции, индекс аккумуляции стока, продольная и поперечная кривизны, топографический индекс влажности (TWI), индекс влажности SAGA, эрозионный (LS) фактор, индекс баланса массы, площадь водосбора, площадь склонового водосбора, вертикальная дистанция до тальвега, глубина долин, относительная позиция на склоне (Olaya, 2004). Для снижения размерности данных о рельефе использован метод главных компонент (Пузаченко, 2004). В результате выделено 4 интегральных латентных переменных – факторы организации рельефа, использованные в качестве предикторов в геоморфологических моделях. Суммарно 4 фактора описывают 82 % дисперсии исходных переменных. Они интерпретированы следующим образом: 1 – принадлежность к положительным или отрицательным формам рельефа (контраст увалов и балок), 2 – расчлененность (контраст платообразного и глубокорасчлененного секторов ландшафта), 3 – экспозиция склона (контраст солнечных и теневых склонов, выраженный через косинус и синус экспозиции), 4 – положение на склоне (контрасты автономных, транзитных и аккумулятивных геохимических позиций). Предметом сравнения для разных сроков были коэффициенты детерминации и стандартизованные регрессионные коэффициенты beta.
Для проверки гипотезы о неодинаковой точности геоморфологических моделей в разных ландшафтных условиях методом однофакторного дисперсионного анализа и его непараметрических аналогов (Kruskal – Wallis) исследовалась локализация остатков моделей, т. е. разности между наблюдаемыми и предсказанными значениями NDVI. В качестве группирующих категоризованных переменных использовались типы фитоценоза, экспозиции склона, типы горных пород, формы рельефа, в качестве зависимых – остатки моделей.
Исходными данными для анализа внутрисезонной динамики фитомассы были приращения значений NDVI между парами сроков в течение года. По стандартизованным (для каждого срока отдельно) значениям приращений и степени их отклонений от модальных приращений определены классы стабильности. Исходное допущение заключалось в следующем. В силу единых для полигона исследования климатических условий за период между любыми двумя сроками в течение вегетационного периода происходит некоторое естественное «фоновое» приращение индекса (с положительным или отрицательным знаком) в большинстве пикселов. Для каждой пары сроков проверялась гипотеза о нормальном распределении приращений или, по крайней мере, наличии модального интервала вблизи среднего значения. Во всех случаях подтвердился второй вариант. Тогда для каждого пиксела можно определить, подчиняется ли он «фоновой» (модальной) тенденции в интервале 0.5 среднеквадратичного отклонения от среднего в каждую сторону либо отклоняется от модального приращения на бóльшую величину (0.5–1.5 и более 1.5 среднеквадратичного отклонения) в положительную или отрицательную сторону. По результатам каждому пикселу присвоен один из пяти «классов отклонения» для данного срока. Затем для всего множества сроков наблюдений проверялась гипотеза о стабильности такого «класса отклонения». Например, пиксел может: а) всегда находиться в модальном классе «фоновых» приращений» (т. е. функционирует «как большинство» пикселов), б) всегда находиться в одном и том же классе сильных отрицательных отклонений (сильно отличается от большинства, но всегда одинаково), в) всегда находиться в классе сильных положительных отклонений, г) часто переходить их одного класса отклонений в другой (иногда «как все», иногда «больше других», иногда «меньше других»). Первые три случая (а, б, в) будут рассматриваться как стабильное функционирование. Последний случай (г) рассматривается как принадлежность пиксела к классу нестабильного функционирования.
Для каждого пиксела рассчитана повторяемость каждого класса отклонений по 13 парам сроков в пределах вегетационного периода в 2014–2019 гг. Для оценки степени стабильности рассчитана мера неопределенности типа динамики по формуле Шеннона. Низкие значения меры неопределенности трактуются как высокая повторяемость одних и тех же классов отклонений (градаций приращения фитомассы), т. е. высокая стабильность сезонного поведения фитоценоза (с вариантами «всегда как большинство», «всегда вразрез с большинством»). Высокая неопределенность трактуется как высокая изменчивость локальной динамики, непостоянство локальных ландшафтных процессов по сравнению с динамикой ландшафтного масштаба, т. е. зависимость от внутрифациальных и межфациальных взаимодействий, сильно варьирующих в зависимости от условий конкретного года или сезона.
На основании классификации пикселов по средним стандартизованным приращениям и среднеквадратическим отклонениям стандартизованных приращений за 13 пар сроков выделены группы динамичности фитомассы, учитывающие как межгодовую, так и внутрисезонную изменчивость.
Для оценки значимости позиционных факторов, отражающих наложение геоциркуляционных и биоциркуляционных структур на геостационарные, для всех пикселов определены расстояния до ближайшей границы урочищ, выделенных на ландшафтной карте (Хорошев, 2016). Методами корреляционного (коэффициент Спирмена) и дисперсионного анализа проверялась гипотеза о том, что неустойчивость фитомассы зависит от близости фации к границам урочищ. Гипотеза о зависимости класса динамики от вклада геоциркуляционных структур внутри урочищ (геостационарных структур) проверялась методами корреляционного и дисперсионного анализа с использованием значений эрозионного (LS) фактора, относительной позиции на склоне, площади водосбора, рассчитанных средствами модуля Terrain Analysis SAGA GIS. Зависимость типов динамики от конфигурации урочищ оценивалась с использованием индекса формы контура (Shape index), который рассчитывался как периметр круга такой же площади, как полигон, деленной на периметр полигона.
Результаты
Полевая верификация классификации космического снимка методом к-средних позволила уверенно распознать следующие классы сообществ: 1) петрофитные степи, 2) сухие типчаковые степи, 3) ковыльно-разнотравные степи, 4) типичные ковыльные степи, 5) луговые степи и луга, 6) кустарники с преобладанием ксерофильных видов, 7) кустарники с преобладанием мезоксерофильных видов, 8) кустарники с преобладанием ксеромезофильных видов с подростом осины, 9) кустарники с преобладанием мезофильных видов с подростом черной ольхи и тополя, 10) черноольшанники и гигрофитные сообщества. В дальнейшем принадлежность к установленным классам использовалась как группирующая переменная в дисперсионных моделях, которые отражают различия степени изменчивости надземной фитомассы между типами фитоценоза.
Измеренные значения фитомассы травяных сообществ после высушивания составили от 5 до 65 ц/га при модальных значениях в интервале 15–25 ц/га. По непараметрическим коэффициентам корреляции Спирмена (К) оценена связь вегетационных индексов с измеренной надземной травяной фитомассой и влажностью почв. Значения NDVI для периода проведения экспедиции (по снимкам Landsat 8 от 06.06.2019 и 22.06.2019) достоверно коррелируют с влажной зеленой фитомассой с одинаковыми К = 0.62. Корреляции с сухой фитомассой ниже, но также достоверны: 0.48 для 06.06.2019 и 0.54 для 22.06.2019. В силу выявленной положительной зависимости ниже в тексте под травяной надземной фитомассой имеется в виду ее оценка по значениям NDVI. Положительная достоверная связь с июньской влажной травяной фитомассой выявлена для NDVI всех сроков летнего периода. Связь весенних NDVI с июньской фитомассой недостоверна.
Установлены различия между видами урочищ по степени адекватности июньской фитомассы как косвенного индикатора накопления фитомассы в другие сезоны года. Для этого результаты полевых измерений в июне 2019 г. сопоставлялись с NDVI соответствующих пикселов, рассчитанными по весенним и летним снимкам Landsat 8 2014–2019 гг. (табл. 1). В урочищах плато и гребней июньская фитомасса отрицательно связана с майскими NDVI, причем максимальные июньские фитомассы с максимальной влажностью фитомассы (плато Актобе) соответствуют минимальным майским NDVI. Минимальные июньские фитомассы, зафиксированные в урочищах узких каменистых гребней, соответствуют максимальным NDVI в мае и апреле, когда эти урочища, в отличие от плато, уже достаточно длительный период находятся в условиях вегетации после освобождения от снежного покрова. В апреле и до середины мая существует отрицательная связь NDVI с июньской влажностью почвы.
Табл. 1. Парные непараметрические коэффициенты корреляции Спирмена, отражающие связь NDVI с полевыми измерениями травяной надземной влажной фитомассы, сухой фитомассы, влажности почв в июне 2019 г.
| Плато и гребни (11 точек) | Склоны (22 точки) | Лощины и балки (18 точек) | |||||||
| Срок измерения NDVI | Влажная фитомасса, ц/га | Сухая фитомасса, ц/га | Влажность гумусового горизонта почв | Влажная фитомасса, ц/га | Сухая фитомасса, ц/га | Влажность гумусового горизонта почв | Влажная фитомасса, ц/га | Сухая фитомасса, ц/га | Влажность гумусового горизонта почв |
| 14.05.2014 | -0.70 | -0.68 | -0.56 | 0.23 | 0.32 | 0.01 | 0.11 | -0.09 | 0.43 |
| 30.05.2014 | 0.59 | 0.69 | 0.56 | 0.43 | 0.46 | 0.00 | 0.35 | 0.17 | 0.62 |
| 08.06.2014 | 0.53 | 0.69 | 0.50 | 0.40 | 0.49 | -0.24 | 0.43 | 0.25 | 0.67 |
| 17.07.2014 | 0.18 | 0.47 | 0.31 | -0.02 | 0.16 | -0.13 | 0.20 | 0.21 | 0.49 |
| 18.06.2015 | 0.32 | 0.54 | 0.29 | 0.52 | 0.58 | -0.05 | 0.36 | 0.20 | 0.50 |
| 13.07.2015 | 0.35 | 0.56 | 0.36 | 0.43 | 0.58 | -0.24 | 0.36 | 0.26 | 0.58 |
| 29.07.2015 | 0.15 | 0.41 | 0.20 | 0.38 | 0.57 | -0.30 | 0.22 | 0.17 | 0.44 |
| 18.05.2018 | -0.79 | -0.82 | -0.94 | 0.13 | 0.34 | -0.14 | 0.11 | 0.01 | 0.34 |
| 25.05.2018 | 0.29 | 0.29 | 0.10 | 0.52 | 0.52 | 0.11 | 0.32 | 0.13 | 0.63 |
| 29.08.2018 | 0.15 | 0.32 | 0.18 | 0.03 | 0.25 | -0.56 | 0.24 | 0.20 | 0.45 |
| 19.04.2019 | -0.92 | -0.84 | -0.77 | -0.26 | -0.27 | -0.08 | -0.10 | 0.10 | -0.24 |
| 05.05.2019 | -0.87 | -0.88 | -0.90 | -0.08 | -0.10 | 0.02 | -0.02 | -0.11 | 0.05 |
| 06.06.2019 | 0.41 | 0.46 | 0.36 | 0.64 | 0.62 | 0.23 | 0.56 | 0.33 | 0.65 |
| 22.06.2019 | 0.58 | 0.73 | 0.56 | 0.49 | 0.52 | -0.04 | 0.58 | 0.34 | 0.64 |
На склонах в период с позднего мая до июля включительно NDVI положительно связан с июньской фитомассой, причем с сухой сильнее, чем с влажной. Корреляция июньской влажности почвы с NDVI любого периода практически отсутствует. В лощинах и балках, наоборот, чем больше июньская влажность почвы, тем выше значения NDVI более поздних летних сроков (см. табл. 1).
Линейные мультирегрессионные геоморфологические модели фитомассы для большинства летних сроков оказались на грани достоверности с коэффициентом детерминации (КД) 0.29–0.35, т. е. с объяснением от 29 до 35 % варьирования. Для большинства майских сроков (кроме позднемайских) модели достоверны с КД 0.22–0.35 и наибольшим вкладом фактора выпуклости, реже – расчлененности и положения на склоне.
Проверка гипотезы об устойчивости во времени видов зависимости вегетационных индексов от факторов дифференциации рельефа (по мультирегрессионным уравнениям) показала следующее (рис. 2). Фактор 1 (контраст положительных и отрицательных форм рельефа) наиболее значим в течение всего года, однако коэффициенты регрессии несколько ниже в весенний период (май), когда обеспеченность почв влагой после снеготаяния достаточно велика как на положительных, так и на отрицательных формах рельефа, а также осенью. В летний период значение фактора заметно возрастает в силу резкой дифференциации почвенной влаги и фитомассы по рельефу: почвы на гребнях и склонах становятся заметно суше, чем почвы лощин и балок, что отражается на больших контрастах фитомассы. Значимость фактора 2 (расчлененности рельефа – контраст останцового плато Актобе и грядово-балочной местности) велика в начале и конце теплого периода. Это свидетельствует о выравнивании условий продуцирования фитомассы между плато и грядово-балочной местностью к концу вегетационного периода. Значимость фактора 3 (экспозиция склона) наиболее велика весной, когда увлажненность почв и фитомассы контрастна между солнечными и теневыми склонами. Значимость фактора 4 (положения в катене – контраст верхних и нижних частей склонов) мала весной, но возрастает в течение летнего периода, что соответствует более сильному обеднению влагой и снижению фитомассы верхних частей склонов по сравнению с нижними частями и подсклоновыми шлейфами.
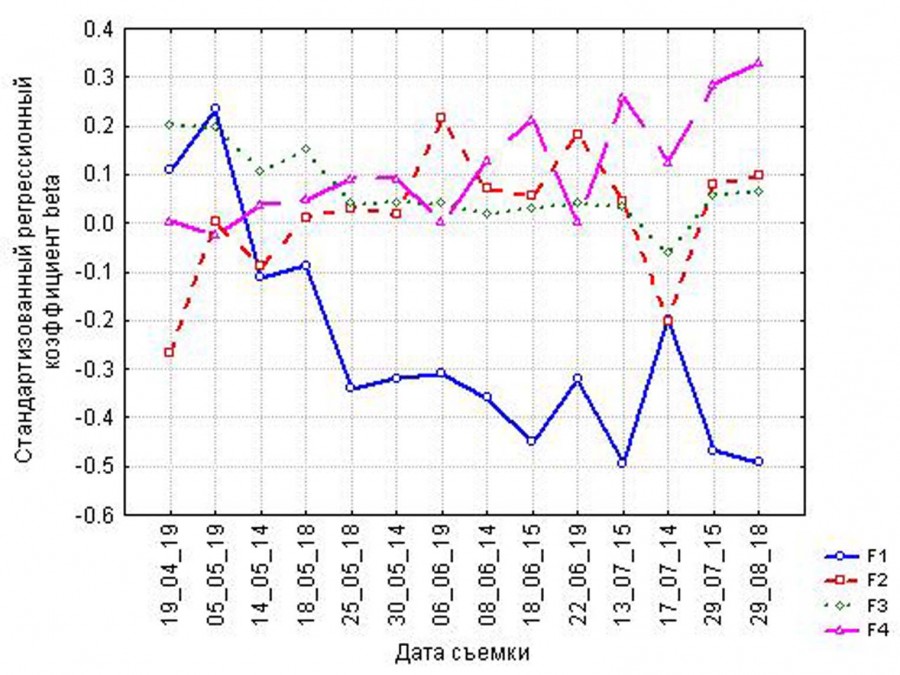
Рис. 2. Динамика вкладов факторов организации рельефа в пространственное варьирование NDVI в течение теплого периода. Стандартизованные регрессионные коэффициенты beta в мультирегрессионном уравнении, где NDVI – зависимая переменная, факторы организации рельефа – независимые переменные (F1 – принадлежность к положительным или отрицательным формам; F2 – расчлененность; F3 – экспозиция склона; F4 – положение на склоне)
Fig. 2. Dynamics of the contributions of the relief organization factors to NDVI spatial variability during the warm period. Standardized regression coefficients beta in the multiple regression equation, where NDVI is the dependent variable, and relief organization factors are the independent variables (F1 – convex or concave landforms; F2 – relief compartmentalization; F3 – slope exposition; F4 – position on the slope)
Анализ отклонений от равновесной и/или стационарной модели, воспроизводимой статистическими методами, представляет особый интерес, т. к. в отклонениях может содержаться важная информация о динамике сообществ (Пузаченко, 2017а). В нашем случае анализ локализации остатков геоморфологических моделей показал неодинаковую тесноту связи фитомассы с рельефом в зависимости от типа фитоценоза, солярной экспозиции склона, геологического строения и разновидности форм рельефа.
Для склоновых урочищ недооценка NDVI геоморфологической моделью характерна для кустарниковых сообществ, особенно в летние сроки (рис. 3). В раннемайские сроки существенной разницы между типами фитоценоза по качеству описания фитомассы геоморфологической моделью нет: остатки модели варьируют примерно в одинаковом диапазоне. В летние месяцы моделью переоценивается фитомасса в типчаковых и ковыльно-разнотравных степях (см. рис. 3). В лощинах в летний период ошибки геоморфологической модели фитомассы зависят от типа фитоценоза. В типчаковых, сухокустарниковых степях реальная фитомасса ниже предсказанной, особенно на крупных монотонных участках. В мезофильных кустарниковых и луговых сообществах реальная фитомасса летом превышает предсказанную.
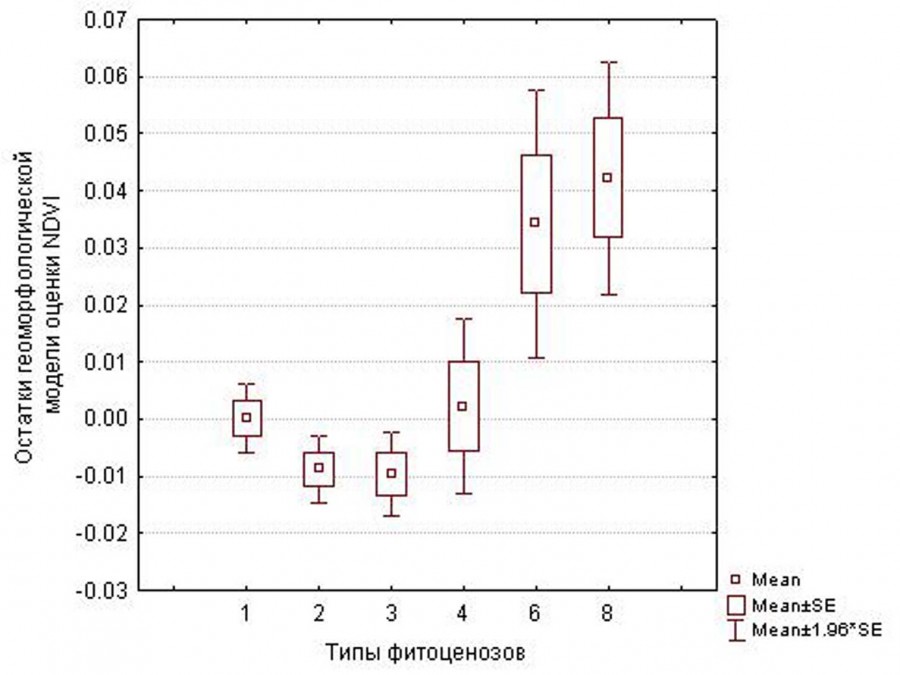
Рис. 3. Диапазоны значений остатков геоморфологических моделей NDVI для 29.07.2015 для типов склоновых фитоценозов: 1 – петрофитные степи, 2 – типчаковые степи, 3 – ковыльно-разнотравные степи, 4 – ковыльные степи, 6 – ксерофильные кустарники, 8 – ксеромезофильные кустарники
Fig. 3. Ranges of residual values in geomorphologic models of NDVI for the types of phytocoenoses on slopes on 29.07.2015: 1 – petrophytic steppes, 2 – Festuca valesiaca dominated steppes, 3 – Stipa and forbs dominated steppes. 4 – Stipa dominated steppes, 6 – xerophilous shrubs, 8 – xeromesophilous shrubs
На склонах южных румбов геоморфологическая модель зеленой фитомассы ландшафтного масштаба завышает реальную фитомассу, особенно в летний период, в меньшей степени – в весенний. Отрицательные остатки модели (фитомасса меньше, чем предсказанная) концентрируются в урочищах с повышенной освещенностью, прогреванием и, следовательно, испарением. Однако для 19.04.2019 хорошо выражена противоположная закономерность: моделью преуменьшается по сравнению с реальностью фитомасса именно прогреваемых южных склонов, где уже начинается вегетация, в отличие от всех остальных.
Перейдем к результатам проверки серии гипотез о зависимости изменчивости NDVI (как суррогата надземной травяной фитомассы) во времени от рельефа и типов фитоценозов.
Общая закономерность внутрисезонной динамики NDVI в степи состоит в наращивании значений к началу лета (соответствующем наращиванию зеленой фитомассы) и постепенному уменьшению к концу лета. Установлены следующие характерные условия рельефа и принадлежность к видам фитоценозов для выделенных групп динамичности фитомассы.
Группа 1 - внутрисезонные приращения существенно ниже фоновых (фитомасса относительно стабильна между сроками в теплый период), но межгодовая вариабельность отклонений велика, т. е. в некоторые годы приращения могут быть существенно выше или ниже фоновых. Такие сообщества встречаются при минимальном эрозионном индексе и максимальной площади водосбора, т. е. по расширенным днищам балок и лощинам в любых типах фитоценозов, но чаще соответствуют приручейным черноольшанникам и зарослям мезофильных кустарников с деревьями.
Группа 2 – внутрисезонные приращения выше фоновых (фитомасса меняется наиболее сильно между сроками), межгодовая вариабельность приращений средняя. Характерна для водосборных понижений, суховатых лощин, краевых частей днищ балок (при минимальном топографическом индексе влажности, максимальном эрозионном факторе), имеет повышенную встречаемость среди луговых степей, ксерофитных склоновых и мезоксерофильных кустарников.
Группа 3 – внутрисезонные приращения чуть ниже фоновых, межгодовая вариабельность приращений средняя. Соответствует плакорам, широким вогнутым понижениям, покатым склонам (в автономных и высоких выпуклых и глубокорасчлененных склоновых позициях) и наиболее характерна для типчаковых степей, реже для ковыльно-разнотравных и ковыльных степей. Для петрофитных степей эта группа наиболее характерна.
Группа 4 - фоновые внутрисезонные приращения с низкой межгодовой вариабельностью; постоянное подчинение фоновой закономерности, общей для всего ландшафта. Соответствует автономным урочищам (с минимальной площадью водосбора и минимальной топографической влажностью), типична для ковыльных степей, в меньшей степени – для типчаковых. Наименее характерна для луговых степей.
Для проверки гипотезы о зависимости внутрисезонной изменчивости NDVI от пространственной конфигурации ландшафтных контуров и положения по отношению к их границам мы воспользовались результатами ландшафтного картографирования.
В автономных урочищах гребней, плато (включая наиболее крупные плато более 40 га) и останцовых холмов при расстоянии от границы урочища более 150 м понижается повторяемость фоновых приращений фитомассы (рис. 4), но повышается повторяемость отрицательных отклонений (быстрое накопление весной и сохранение летом). При этом повышена неопределенность динамических состояний, т. е. стабильность динамики невысокая. Однако если исключить из расчета урочища крупных плато с площадью более 40 га, то по мере удаления от границ автономных урочищ повторяемость отклонений от фоновой динамики, наоборот, снижается, а фоновой динамики – растет, как и в целом стабильность. Аналогичная закономерность выявлена для трансаккумулятивных урочищ подсклоновых шлейфов и конусов выноса: на расстоянии более 100 м от границ снижается повторяемость фоновых внутрисезонных приращений NDVI и увеличивается нестабильность динамики.
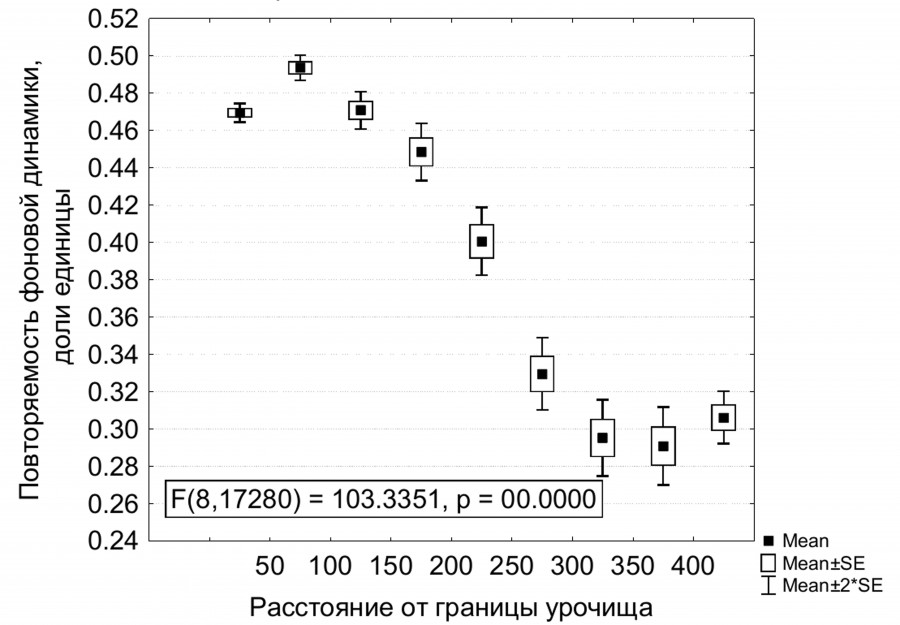
Рис. 4. Повторяемость фоновых внутрисезонных приращений фитомассы (по 13 парам сроков) в зависимости от расстояния до границы для автономных плакорных степных урочищ
Fig. 4. Repetition of background intra-seasonal increments of phytomass (for 13 pairs of terms) depending on the distance to the boundary for the autonomous plakor steppe stows
Наибольшую площадь на территории (68 %) составляют склоны с крутизной более 3°. В группе склоновых урочищ расстояние до границы не является фактором стабильности биомассы. Наибольшая повторяемость фоновых приращений фитомассы и наибольшая стабильность характерны для верхних частей склонов при минимальном значении эрозионного (LS) фактора (рис. 5). По мере возрастания площади водосбора в нижней части склонов внутрисезонная изменчивость и разнообразие вариантов динамики фитомассы возрастают.
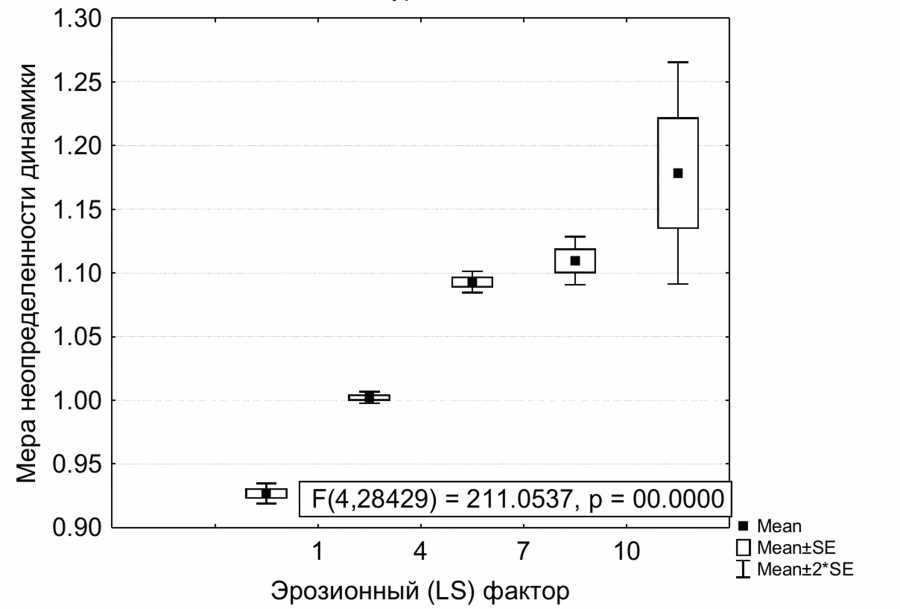
Рис. 5. Зависимость меры неопределенности (индекс Шеннона) внутрисезонной динамики NDVI от эрозионного фактора рельефа для склоновых урочищ
Fig. 5. Dependence of the uncertainty measure (Shennon index) of NDVI intra-seasonal dynamics on the erosion factor for the slope stows
Для автономных урочищ (гребни, плато, останцовые холмы) установлена зависимость изменчивости внутрисезонных приращений фитомассы от конфигурации урочища, оцененной по индексу формы (shape index). В срединном диапазоне значений индекса формы имеет место следующая зависимость: чем более компактна форма урочища (малые значения индекса формы), тем ниже повторяемость фоновых приращений (рис. 6). Максимальная повторяемость фоновой динамики свойственна самым вытянутым автономным урочищам узких гребней.
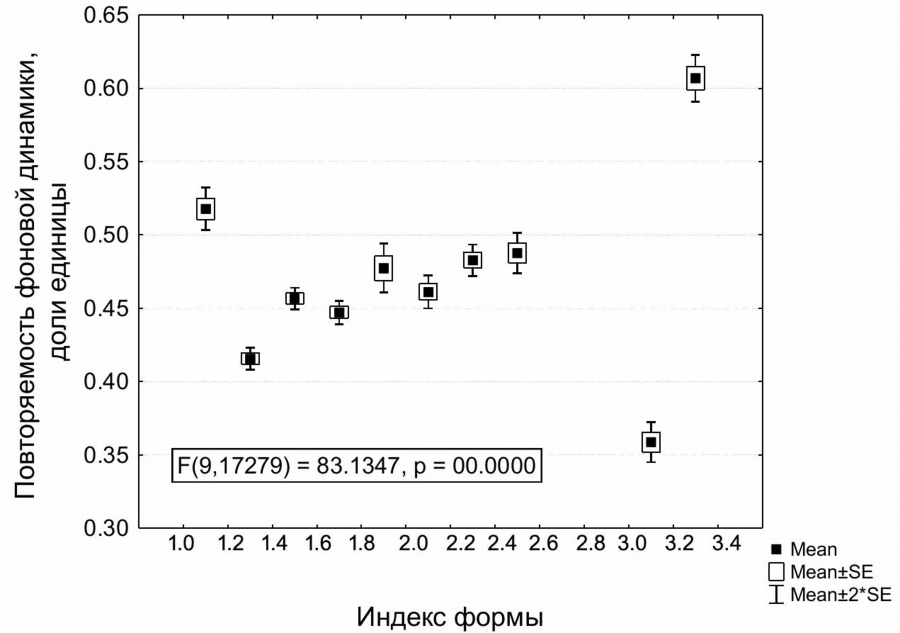
Рис. 6. Повторяемость фоновых внутрисезонных приращений фитомассы (по 13 парам сроков) в зависимости от индекса формы для автономных плакорных степных урочищ
Fig. 6. Repetition of background intra-seasonal invrements of phytomass (for 13 pairs of terms) depending on the shape index for the autonomous plakor steppe stows
Обсуждение
Установленные положительные достоверные корреляции между NDVI и полевыми измерениями дают основания рассматривать NDVI, с некоторыми оговорками, как количественный индикатор зеленой травяной фитомассы. Очевидно, что NDVI отражает не только травяную, но и кустарниковую фитомассу, которая не попадала в укос. Поэтому неслучайно, что основные отклонения от прямой зависимости связаны со значениями сырой травяной надземной фитомассы более 60 ц/га, которая наблюдается в сильно закустаренных лощинах и балках. В автономных и трансэлювиальных типичностепных и петрофитно-степных урочищах связь между зеленой фитомассой и NDVI положительная и практически линейная. Это подтверждает рекомендацию, отраженную в литературе, что для использования NDVI как суррогата зеленой фитомассы необходимо предварительно строго устанавливать диапазон условий, для которых существует статистически значимая связь, а также учитывать межгодовое варьирование тесноты связи (Doraiswamy et al., 2005; Raynolds et al., 2006; Araya et al., 2017; Nagy et al., 2018).
Фитомасса с начала июня и до конца лета меняется примерно пропорционально на большей части территории. В то же время весенние условия формирования фитомассы принципиально иные, по сравнению с июньскими, за счет влияния длительности снеготаяния, экспозиции склона, разного начала вегетации в разных урочищах. Сохранение максимального запаса почвенной влаги в июне в черноземах на плато Актобе и соответствующий максимум фитомассы следуют за сильной весенней задержкой развития фитомассы. Максимальная весенняя фитомасса развивается на гребнях, которые наиболее рано прогреваются и освобождаются от снега, а к июню влажность почвы резко снижается и начинается летний перерыв вегетации большинства злаков. На склонах имеет место «разрыв» между условиями биопродуцирования раннего лета, связанными с наследием снеговой влаги, и более поздних периодов, когда фитомасса в основном определяется дождями. В лощинах и балках режимов биопродуцирования в течение лета унаследованы от почвенной влаги, накопленной главным образом за счет снеготаяния.
Анализ остатков «геоморфологической модели» надземной фитомассы, оцененной по NDVI, показал, что в наиболее развитых увлажненных фациях лощин и балок летом запас фитомассы существенно выходит за рамки, предопределенные потоками влаги по рельефу. Это свидетельствует о наличии механизмов положительной обратной связи, когда хорошо развитая почва (чернозем гидрометаморфизованный или выщелоченный) стимулирует развитие фитомассы и наоборот. Отметим, что именно в днищах лощин и балок NDVI, рассчитанные по снимкам 06.06.2019 и 22.06.2019, наиболее сильно отклоняются в сторону завышения от значений, предсказываемых регрессионной моделью по измеренным в поле значениям травяной фитомассы. Это связано, безусловно, с большим вкладом зеленой массы кустарников в этих видах урочищ. На склонах остатки «геоморфологической модели» в значительной степени контролируются экспозицией. На солнечных склонах фитомасса в период вегетации сокращается в большей степени, чем на теневых, по сравнению со значениями, предопределенными условиями рельефа.
Исследование показало существенные отличия типов динамики фитомассы в краевых и внутренних секторах автономных плакорных урочищ. В первом случае преобладает внутрисезонная динамика фонового типа, во втором – сильная изменчивость типов динамики. Отличительная особенность внутренних частей плато – развитие хорошо развитых черноземов южных с мощностью гумусированного профиля (А + АВ) 27–37 см под зональными житняково-мятликово-тонконогово-залесскоковыльными степями. Возможно, благодаря высокому видовому богатству и стабильности морфолитогенной основы в зависимости от условий конкретного периода создаются конкурентные преимущества разных групп видов, что и дает в результате нестабильное поведение по сравнению с фоновым ландшафтом. Следовательно, в центральных секторах таких урочищ снижается зависимость от обычной внутрисезонной погодной динамики ландшафтного масштаба, но при этом включаются механизмы саморегулирования, связанные с положительными обратными связями в системе «почва – фитоценоз», т. е. формируется локальная пространственно-временная устойчивость целостных экосистем (Пузаченко, 2017б). Урочища плато «живут самостоятельной жизнью» по сравнению с остальной территорией. При этом возможно нескольких устойчивых модулей, т. е. проявление пластичности (Пузаченко, 2017а). На периферии плато по мере сокращения мощности гумусового профиля и перехода преобладания к овсецово-лессингоковыльным степям повторяемость фоновой динамики растет. Итак, на плато, внутри геостационарной единицы с одинаковым рельефом и геологическим строением обособляются разные биоциркуляционные структуры. Параллельно проявляется и зависимость от конфигурации урочища. Причина повышенной нестабильности продуцирования биомассы в крупных компактных урочищах видится в ослаблении латеральных взаимодействий и возможности усиления радиальных почвенно-фитоценотических положительных обратных связей. При этом нельзя сказать, что динамика фитомассы становится более стабильной: локальные факторы могут вызывать самые разнообразные отклонения от фоновой динамики, что отражается на повышенных значениях меры неопределенности типов внутрисезонной динамики. При вытянутой или изрезанной форме возрастает интенсивность латеральных взаимодействий с соседними урочищами, т. е. взаимодействий, порождающих собственно ландшафт как систему пространственных элементов. Именно при таких условиях наиболее часто проявляется фоновая динамика NDVI, отражающая единообразную реакцию степной фитомассы на внутрисезонные погодные изменения, – рост оптимальности увлажнения весной, снижение влагообеспеченности в течение лета и т. п. В то же время из этой зависимости выпадают урочища с максимальными и, наоборот, очень низкими значениями индекса формы.
В группе склоновых урочищ доказано наличие внутриурочищного варьирования типов внутрисезонной динамики фитомассы. В терминологии ландшафтоведения это означает, что геоциркуляционная организация на склонах более существенна для временнóй организации, чем геостационарная: при прочих равных геолого-геоморфологических условиях миграция влаги вниз по склону создает различия в характере функционирования.
Заключение
В низкогорно-степном ландшафте Айтуарской степи вегетационный индекс NDVI адекватно отражает надземную травяную фитомассу начала летнего периода за исключением наиболее высокопродуктивных луговых и лугово-степных сообществ с большим участием кустарников. Характер зависимости фитомассы от ландшафтных условий варьирует в пространстве. Фитомасса в лощинах и балках в течение всего лета зависит от почвенной влаги, имеющейся в начале лета и накопленной в период весеннего снеготаяния, в то время как на склонах фитомасса зависит в основном от летних дождей без связи с раннелетней влажностью почвы.
Вид зависимости травяной фитомассы от факторов организации рельефа меняется в течение теплого периода. Вклад факторов принадлежности к положительным или отрицательным формам рельефа и положения в катене возрастает в летний период по сравнению с весенним, что отражает увеличение контрастов условий почвенного увлажнения. Факторы расчлененности рельефа и инсоляции более значимы в поздневесенний период из-за разной скорости истощения снеговой влаги в почвах на плато и в грядово-балочной местности, на солнечных и теневых склонах.
Типы внутрисезонной динамики фитомассы выражены как разные варианты подчиненности растительных сообществ фоновым погодным изменениям ландшафтного масштаба и локальным факторам. Ландшафтные единицы ранга урочищ, обусловленные геостационарными структурами, могут иметь существенные внутренние различия по типам внутрисезонной динамики продуцирования фитомассы в зависимости от водосборной площади и близости к границам. Существенное варьирование типов динамики (нестабильность) характерно для центральных секторов крупных степных урочищ плато и подсклоновых шлейфов, видимо, в силу возрастания значимости локальных почвенно-фитоценотических обратных связей. В нижних частях склонов типы динамики сильно варьируют из-за нестабильности почвенного увлажнения. Стабильная подчиненность фоновой внутрисезонной динамике возникает при активной вовлеченности в латеральные взаимодействия с соседними урочищами, что характерно для урочищ с малой компактностью, особенно для гребней.
В целом исследование показало необходимость сопряженного учета геостационарных, геоциркуляционных и биоциркуляционных ландшафтных структур при объяснении временной изменчивости травяной надземной фитомассы.
Библиография
Гродзинський М. Д. Ландшафтна екологія . К.: Знання, 2015. 550 с.
Пахучий В. В., Пахучая Л. М. Опыт использования вегетационных индексов при комплексных исследованиях на объектах гидролесомелиорации // Вестник Поволжского государственного технологического университета. Серия: Лес. Экология. Природопользование. 2014. № 1 (21). С. 33–41.
Пузаченко Ю. Г. Биогеоценоз как сложная динамическая система // Биогеоценология в XXI веке: идеи и технологии. М.: Товарищество научных изданий КМК, 2017а. С. 11–114.
Пузаченко Ю. Г. Математические методы в экологических и географических исследованиях . М.: Академия, 2004. 416 с.
Пузаченко Ю. Г. Организация ландшафта // Вопросы географии. Сб. 138. Горизонты ландшафтоведения. М.: Кодекс, 2014. С. 35–64.
Пузаченко Ю. Г. Теоретико-методологические основы долговременных эколого-географических исследований на территории заповедников // Вопросы географии. Сб. 143. Эколого-географические исследования на заповедных территориях. М.: РГО, 2017б. С. 192–233.
Современное ландшафтно-экологическое состояние и проблемы оптимизации природной среды регионов: Материалы XIII Международной ландшафтной конференции, посвященной столетию со дня рождения Ф. Н. Милькова . Воронеж: ИСТОКИ, 2018. Т. 1. 489 с. Т. 2. 426 с.
Солнцев В. Н. Структурное ландшафтоведение: основы концепции. Некоторые аргументы . М., 1997. 12 с.
Хорошев А. В. Ландшафтная структура Айтуарской степи (заповедник «Оренбургский») и экологические ряды урочищ // Проблемы географии Урала и сопредельных территорий: Материалы IV Всероссийской научно-практической конференции с международным участием. Челябинск: Край Ра, 2016. С. 210–216.
Хорошев А. В., Леонова Г. М. Реакции при изменении увлажнения в ландшафте Айтуарской степи (Южный Урал) // Вестник Московского университета. Серия 5: География. 2015. № 4. С. 95–103.
Araya S., Ostendorf B., Lyle G., Lewis M. Remote Sensing Derived Phenological Metrics to Assess the Spatio-Temporal Growth Variability in Cropping Fields // Advances in Remote Sensing. 2017. Vol. 6. P. 212–228. DOI: 10.4236/ars.2017.63016.
Basso B., Ritchie J. T., Pierce F. J., Braga R. P. and Jones J. W. Spatial validation of crop models for precision agriculture // Agricultural Systems. 2001. Vol. 68. P. 97–112. DOI: 10.1016/S0308-521X(00)00063-9.
Corwin D. Site-specific management and delineating management zones // Oliver M. (Ed.) Precision Agriculture for Food Security and Environmental Protection. Ear5thscan. London, UK, 2013. P. 135–157.
Doraiswamy P. C., Sinclair T. R., Hollinger S., Akhmedov B., Stern A. and Prueger J. Application of MODIS Derived Parameters for Regional Crop Yield Assessment // Remote Sensing of Environment. 2005. Vol. 97. P. 192–202. DOI: 10.1016/j.rse.2005.03.015.
Kaspar T. C., Colvin T. S., Jaynes D. B., Karlen D. L., James D. E., Meek D. W., Pulido D., Butler H. Relationship between six years of corn yields and terrain attributes // Precision Agriculture. 2003. Vol. 4. P. 87–101. DOI: 10.1023/A:1021867123125.
Nagy A., Fehér J., Tamás J. Wheat and maize yield forecasting for the Tisza river catchment using MODIS NDVI time series and reported crop statistics // Computers and Electronics in Agriculture. 2018. Vol. 151. P. 41–49. DOI: 10.1016/j.compag.2018.05.035.
Olaya V. A gentle introduction to SAGA GIS. Edition 1.2. 2004. 202 p.
Raynolds M. K., Walker D. A., Maier H. A. NDVI patterns and phytomass distribution in the circumpolar Arctic // Remote Sensing of Environment. 2006. Vol. 102. P. 271–281. DOI: 10.1016/j.rse.2006.02.016.
Turner M. G. & Gardner R. H. Landscape ecology in theory and practice. Pattern and process. N. Y.: Springer, 2015. 482 p.
Verhulst N., Govaerts B. The normalized difference vegetation index (NDVI) GreenSeekerTM handheld sensor: Toward the integrated evaluation of crop management. Part A: Concepts and case studies. Mexico, D. F.: CIMMYT, 2010. P. 1–16.
Благодарности
Исследование выполнено при финансовой поддержке Российского фонда фундаментальных исследований (проект 20-05-00464).

 © 2011 - 2024
© 2011 - 2024