



Введение
Скорость роста и продолжительность жизни лишайников зависят от генетических особенностей конкретного вида и от характеристик места обитания, в особенности: климатических условий, химического состава субстрата и физических свойств субстрата (Викторов, 1956; 1960; Домбровская, 1963; Innes, 1985; Галанин, 1997). Работы по изучению роста разных экологических групп лишайников активно проводятся с середины ΧΧ столетия. За этот период в работах ряда авторов (Platt, Amsler, 1955; Beschel, 1960; Armstrong, 1974) выделяются фазы роста эпилитных лишайников (Armstrong, 1974): 1) долинейная (прирост с возрастом нарастает в логарифмической прогрессии); 2) линейная (ежегодный прирост постоянный); 3) постлинейная (ежегодный прирост постепенно уменьшается).
Самая медленная скорость роста отмечена у накипных лишайников и составляет 0.01…0.70 мм/год (Andrews, Weber, 1964; Турманина, 1979). В связи с этим, по оценкам исследователей, накипные эпилитные лишайники могут иметь значительно большую продолжительность жизни по сравнению с другими биоморфами лишайников или другими организмами. Так, в циркумполярных и высокогорных областях продолжительность жизни накипных жизненных форм лишайников может достигать нескольких тысяч лет (Beshel, 1957; Reger, Pewe, 1969; Calkin, Ellis, 1980). Возраст отдельных талломов лишайника Rizocarpon geographicum (L.) DC. в полярных районах составляет 4000…4500 лет (Beschel, 1960), по другим данным, 8500…9000 лет (Denton, Karlen, 1973; Miller, Andrews, 1972). В умеренной зоне средняя продолжительность жизни накипных видов лишайников значительно ниже — 500…600 лет (Галанин, Глушкова, 2003; Галанин, 2012).
Изучение особенностей роста и определение возраста эпилитных лишайников являются актуальными и недостаточно изученными вопросами в лихенологии. Большой интерес представляет практическое применение знаний о росте эпилитных лишайников, в частности, в лихенометрическом датировании археологических и геологических объектов с помощью видов лишайников, обладающих большой продолжительностью жизни (например, Rhizocarpon geograficum). Метод лихенометрического датирования обладает технической простотой и низкой стоимостью. Однако для его применения нужно проводить подбор лишайников-индикаторов, изучать рост вида-индикатора в конкретных физико-климатических условиях и решать методические проблемы точности и воспроизводимости датировок (Галанин, Глушкова, 2003).
О росте эпилитных лишайников для территории Карелии нет данных. Поэтому основной целью настоящего исследования является изучение особенностей роста эпилитных лишайников в условиях Карелии на примере вида Protoparmeliopsis muralis (Schreb.) M. Choisy и оценка возраста талломов. Также на основании проведенного исследования роста Protoparmeliopsis muralis оценена возможность использования вида в лихенометрическом датировании.
Материалы
Protoparmeliopsis muralis является облигатным эпилитом, характеризуется плагиотропным типом роста, имеет радиальный таллом, который относится к классу накипных диморфных: центральная часть представлена чешуйчатым морфотипом, а краевая часть – лопастным (Purvis at al., 1992).
Исследование проводилось на территории Государственного природного заповедника «Кивач» (Кондопожский район, Республика Карелия) в условиях среднетаежной подзоны. Изучение роста вида выполнено на мониторинговой площадке, заложенной в 2007 г. для изучения роста нескольких видов эпилитных лишайников, на диабазовых скальных выходах в условиях супралиторали побережья реки Суны. Исследованный участок не подвергается прямому воздействию со стороны реки (рис. 1).
В течение 11 лет (2007, 2008, 2011, 2012, 2013 2017 и 2018 гг.) исследовался рост 17 талломов Protoparmeliopsis muralis. За период исследования проведено 117 измерений площадей талломов и получено 57 значений годовых приростов.

Рис. 1. Мониторинговая площадка изучения роста Protoparmeliopsis muralis
Fig. 1. Monitoring plot for study of growth of Protoparmeliopsis muralis
Методы
Для изучения роста талломов был использован метод картирования (Фадеева, Сонина, 2000, 2001). В полевых условиях в сухую солнечную погоду (не менее трех солнечных дней до момента измерения) на прозрачную полиэтиленовую пленку обводились контуры талломов с использованием лупы (х7-кратное увеличение). В лабораторных условиях контуры сканировалась вместе с линейкой, по шкале линейки проводилось масштабирование и измерение площади талломов в программе Autocad. В качестве индекса возраста таллома использован показатель – площадь таллома (см2). Для создания моделей роста Protoparmeliopsis muralis применен регрессионный анализ в среде Excel (Ивантер, Коросов, 2003). Возраст талломов оценивался по моделям роста.
Результаты
У исследуемых талломов Protoparmeliopsis muralis площадь варьирует от 1 до 26 см2, годовые приросты изменяются в диапазоне от 0.04 до 1.39 см2. Распределение размеров талломов имеет резкую правостороннюю асимметрию (коэффициент асимметрии – A = 0.818, ошибки коэффициента асимметрии – mA = 0.224, фактический и теоретический коэффициенты Стьюдента – ТА = 3.66 > Tтабл. = 1.96). Наибольшая доля вариант (35 %) сосредоточена в первом классе вариационного ряда (1…4 см2). Основная масса вариант (74 %) находится в первых 3 классах (1…12 см2), в остальных соответственно – 26 % вариант. Заметный провал частот наблюдается в 6-м и 7-м классах вариационного ряда площадей талломов, в которые попали талломы максимальных размеров с площадью 21…26 см2 – 4 % талломов (табл. 1).
Таблица 1. Распределение площадей талломов Protoparmeliopsis muralis, см2
Table 1. Distribution of areas of thallii Protoparmeliopsis muralis, cm2
| Классы | Классовые интервалы, см2 | Центр класса, см2 | Частота, талломов | Доля, % | Накопленные доли, % |
| 1 | 1…4 | 2.50 | 41 | 35.04 | 35.04 |
| 2 | 5…8 | 6.50 | 26 | 22.22 | 57.26 |
| 3 | 9…12 | 10.50 | 19 | 16.24 | 73.50 |
| 4 | 13…16 | 14.50 | 15 | 12.82 | 86.32 |
| 5 | 17…20 | 18.50 | 12 | 10.26 | 96.58 |
| 6 | 21…24 | 22.50 | 2 | 1.71 | 98.29 |
| 7 | 25…28 | 26.50 | 2 | 1.71 | 100.00 |
При логарифмировании значений площадей талломов Protoparmeliopsis muralis распределение становится нормальным, это свидетельствует о том, что размеры талломов имеют логнормальное распределение (критерий Колмогорова – Смирнова λ = 0.63 < 1.36(a = 0.05)). Вероятность появления значений в интервале 1.17…26.39 см2 для логнормального распределения составляет 94 %. Мода, медиана и среднее арифметическое (табл. 2), рассчитанные для логнормального распределения, хорошо описывают фактическое распределение признака, представленное в табл. 1.
Таблица 2. Площадь талломов Protoparmeliopsis muralis, см2
Table 2. Area of thallii Protoparmeliopsis muralis, cm2
| Min | Max | n | Mo | Me | µ | S2 |
| 1.17 | 26.39 | 117 | 3.61 | 6.78 | 9.29 | 75.64 |
Примечание. Min – минимальное значение; Max – максимальное значение; n – количество вариант; Mo – мода; Me – медиана; µ – математическое ожидание; S2 – дисперсия.
Распределение годовых приростов талломов также имеет резкую правостороннюю асимметрию и значимо отличается от нормального распределения (коэффициент асимметрии – A = 0.653, ошибки коэффициента асимметрии – mA = 0.316, фактический и теоретический коэффициенты Стьюдента – ТA = 2.065 > T (0.05,∞) = 1.96). Наибольшая частота и доля признака смещены влево и находятся во 2-м классе вариационного ряда (0.25…0.45 см2 в год, 28 %).
Основная масса вариант (68 %) находится в диапазоне первых трех классов (0.04…0.66 см2 в год). В остальных классах (4…7) сосредоточенно 32 % вариант, имеющих значения в пределах 0.67…1.39 см2 (табл. 3). Логарифмирование годовых приростов также приводит к логнормальному распределению (критерий Колмогорова – Смирнова λ = 0.28 < 1.36(a = 0.05)). С вероятностью 93 % годовые приросты будут находиться в интервале 0.14…1.39 см2 в год. Мода, медиана, среднее, дисперсия, стандартное отклонение (табл. 4), рассчитанные для логнормального распределения годовых приростов, хорошо описывают фактическое распределение признака, представленное в табл. 3. Годовые приросты в диапазоне 0.04…0.10 см2 (7 значений) находятся в пределах погрешности измерения контуров талломов (0.1 см2), также эти значения значительно ниже модельных скоростей роста талломов (рис. 4) и в расчете статистик логнормального распределения не учитывались.
Таблица 3. Распределение годовых приростов Protoparmeliopsis muralis, см2 в год
Table 3. Distribution of annual growth of thallii Protoparmeliopsis muralis, cm2 per year
| Классы | Классовые интервалы, см2 | Центр класса, см2 | Частота, талломов | Доля, % | Накопленные доли, % |
| 1 | 0.04…0.24 | 0.14 | 12 | 21.05 | 21.05 |
| 2 | 0.25…0.45 | 0.35 | 16 | 28.07 | 49.12 |
| 3 | 0.46…0.66 | 0.56 | 11 | 19.30 | 68.42 |
| 4 | 0.67…0.87 | 0.77 | 5 | 8.77 | 77.19 |
| 5 | 0.88…1.08 | 0.98 | 5 | 8.77 | 85.96 |
| 6 | 1.09…1.29 | 1.19 | 6 | 10.53 | 96.49 |
| 7 | 1.30…1.50 | 1.40 | 2 | 3.51 | 100.00 |
Таблица 4. Годовые приросты талломов Protoparmeliopsis muralis, см2 в год
Table 4. Annual growth of thallii Protoparmeliopsis muralis, cm2 per year
| Min | Max | Мo | Me | µ | S2 | n |
| 0.14 | 1.39 | 0.35 | 0.52 | 0.63 | 0.18 | 50 |
На основании данных об изменении размеров талломов по годам созданы модели роста каждого таллома за период исследования (11 лет). На рис. 2 представлены модели роста некоторых исследуемых талломов. У всех талломов в диапазоне размеров 1…26 см2 зафиксирован линейный рост, модели обладают высокими коэффициентами аппроксимации.
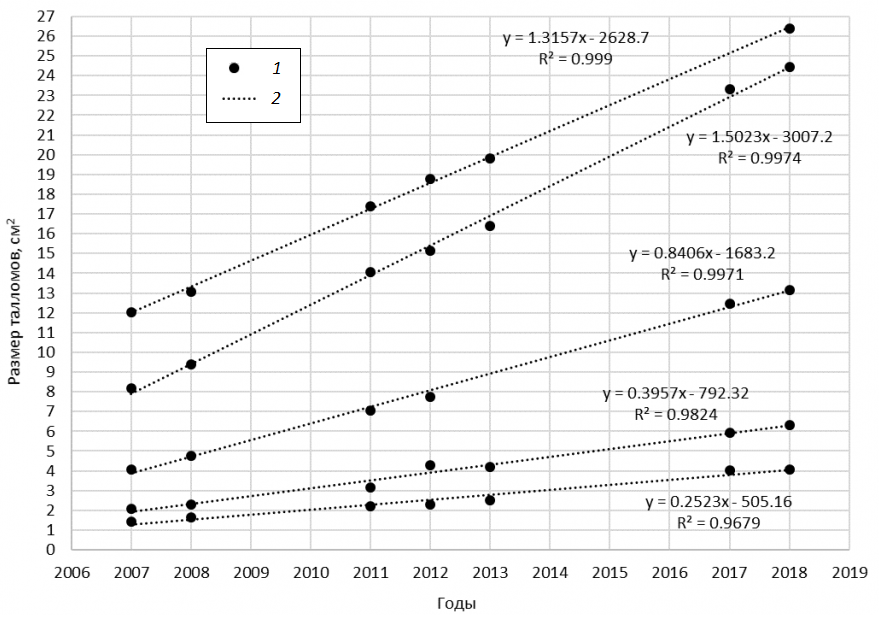
Рис. 2. Рост талломов Protoparmeliopsis muralis за период исследования: 1 – эмпирические данные, 2 – линейный тренд, R2 – коэффициент аппроксимации
Fig. 2. Growth of Protoparmeliopsis muralis over the study period: 1 – empirical data, 2 – linear trend, R2 – coefficient of approximation
Так как талломы данного вида растут линейно, то и скорость роста талломов в моделях постоянна, что отражено горизонтальными линиями на рис. 3. Скорость роста варьирует в пределах 0.20…1.50 см2 в год.
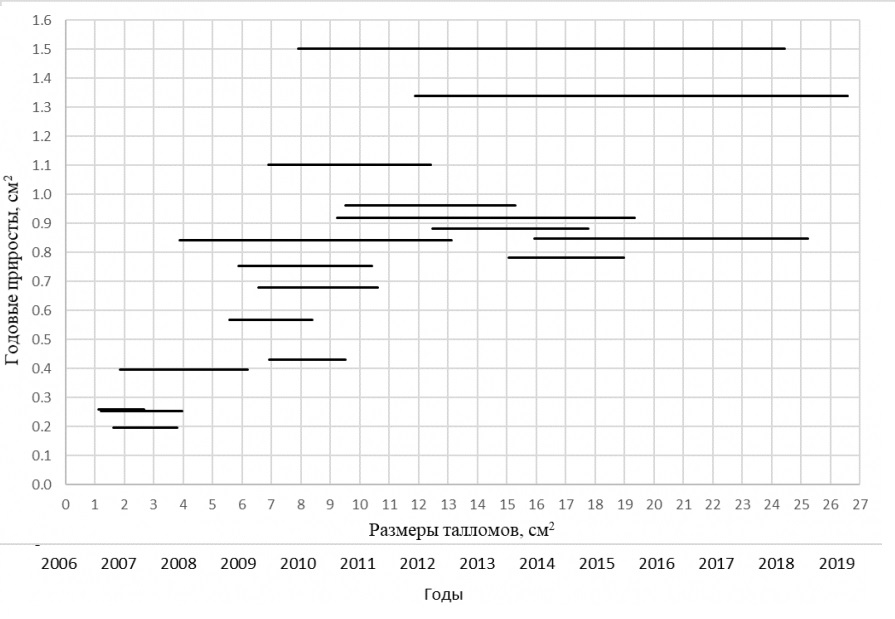
Рис. 3. Скорость роста Protoparmeliopsis muralis, см2 в год
Fig. 3. Growth rate of Protoparmeliopsis muralis, cm2 per year
Учитывая, что выявленный линейный рост Protoparmeliopsis muralis зафиксирован для талломов в интервале 1…26 см2, модели, оценивающие продолжительность линейного роста (рис. 4), построены с учетом этого диапазона. Продолжительность линейного роста в моделях составляет 6…23 года.
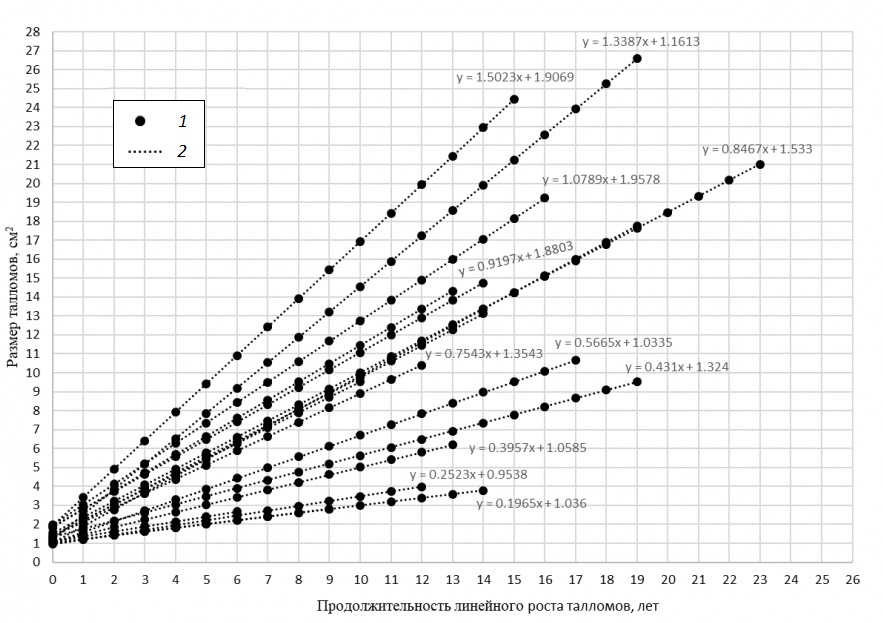
Рис. 4. Оценка времени роста талломов в диапазоне размеров 1…26 см2: 1 – модельные значения, 2 – линейный тренд
Fig. 4. Estimation of growth time of the Protoparmeliopsis muralis in the size range 1...26 cm2: 1 – model values, 2 – linear trend
Распределение продолжительности роста в выборке можно считать нормальным (A = 0.07, mA = 0.55, TA = 0.12 < Tтабл. = 1.96), эксцесс значимо от нормального не отличается (E = 0.19, mE = 1.06, TE = 0.12 < Tтабл. = 1.96). Большая часть исследуемых талломов находится в 3-м классе вариационного ряда (35.29 %) и имеет продолжительность роста 14…17 лет (табл. 5).
Таблица 5. Распределение продолжительности линейного роста Protoparmeliopsis muralis
Table 5. Distribution of linear growth duration of the Protoparmeliopsis muralis
| Классы | Классовые интервалы, см2 | Частота, талломов | Доля, % | Накопленные доли, % |
| 1 | 6…9 | 2 | 11.76 | 11.76 |
| 2 | 10…13 | 5 | 29.41 | 41.18 |
| 3 | 14…17 | 6 | 35.29 | 76.47 |
| 4 | 18…21 | 3 | 17.65 | 94.12 |
| 5 | 22…25 | 1 | 5.88 | 100.00 |
Средняя продолжительность роста талломов составляет 14 лет (табл. 6), доверительный интервал средней составляет 12.25…16.57 года (P = 0.95, a = 0.05, df = 16).
Таблица 6. Продолжительность линейного роста Protoparmeliopsis muralis
Table 6. Linear growth duration of Protoparmeliopsis muralis
| Min | Max | М | mM | S | mS | CV | n |
| 6 | 23 | 14.41 | 1.02 | 4.20 | 0.72 | 29.14 | 17 |
Примечание. М – средняя арифметическая, S – стандартное отклонение, mM – ошибка средней арифметической, mS – ошибка стандартного отклонения, CV – коэффициент вариации.
Время роста распределяется по нормальному закону, это позволяет прогнозировать максимальную продолжительность роста Protoparmeliopsis muralis в заданном диапазоне размеров с помощью интервальной оценки. Так, в соответствии с правилом 3 сигм (для нормального распределения) признак редко отклоняется от среднего значения больше чем на 3 стандартных отклонения. При доверительной вероятности P = 0.999, уровне значимости а = 0.001, числе степеней свободы df = 16, 99.9 % талломов будет находиться в интервале M ± 4.015⋅S, при этом можно ожидать появление максимального времени роста 29 лет.
Обсуждение
Полученные в ходе исследования данные о росте в пределах небольшого скального фрагмента свидетельствуют о большом разбросе значений размеров талломов (от 1 до 26 см2) и их модельных приростов (от 0.2 до 1.50 см2). Скорость роста талломов Protoparmeliopsis muralis в полученных моделях отличается в 7.5 раза. Можно предположить, что максимальные размеры в ценопопуляции будут иметь талломы, обладающие высокой скоростью роста, талломы с низкой скоростью роста должны иметь меньшие размеры в силу ограниченной продолжительности жизни организмов. Поэтому самый большой таллом в выборке не обязательно будет самым старым, а значительно различающиеся талломы по размерам в выборке могут иметь примерно одинаковый возраст. Например, в моделях продолжительность линейного роста самого большого таллома, достигшего 26 см2, имеющего скорость роста 1.34 см2 в год, составляет 19 лет, а таллома, достигшего 21 см2 и имеющего скорость роста 0.85 см2 в год, 23 года, и он является самым старым в выборке. Это отмечается и в литературе, например, в исследованиях М. Бешеля (Beschel, 1960) указывается, что размеры талломов Rizocarpon geographicum в возрасте 100 лет (лишайниковый фактор) варьируют от 2 до 45 мм (приросты 0.02…0.60 мм/г) в зависимости от экологических условий.
Выявленное резко правосторонне асимметричное распределения размеров талломов (площадей) в вариационном ряду у Protoparmeliopsis muralis согласуется с данными по размерам талломов (измеренные поперечники) для эпилитных видов Caloplaca sp., Xanthoria sp. (Галанина, 2011), такое же распределение указывается и для листоватых эпифитных лишайников Lobaria pulmonaria (L.) Hoffm (Михайлова, 2005) и Physconia detersa (Nyl.) Poelt. (Галанина, 2011). По данным литературы, распределения размеров талломов могут отражать стадии развития локальной популяции эпилитных лишайников. Распределения значений признака, близкие к нормальному, указывают на то, что колонизация поверхности субстрата закончилась. Распределения с модой в области минимальных значений свидетельствуют о том, что колонизация поверхности продолжается, маленькие подселяющиеся талломы увеличивают частоты их встречаесмости в области минимальных значений, вызывая правую асимметрию распределения, или что старые талломы в результате конкуренции разрушаются. Распределения с правосторонней асимметрией свидетельствуют об уменьшение количества молодых талломов в ценопопуляции (Farrar, 1974).
Кроме того, по типу распределения можно предсказать размах признаков, например размеров талломов, их годовых приростов и продолжительности роста, и отбраковывать аномальные максимальные и минимальные значения этих параметров из выборки. Вероятность появления значений в интервале площадей талломов 1.17…26.39 см2 для логнормального распределения составляет 94 %, с вероятностью 93 % годовые приросты будут находиться в интервале 0.14…1.39 см2 в год. Так, полученные фактические максимальные значения размеров и годовых приростов Protoparmeliopsis muralis не превышают вероятности 95 %, это может свидетельствовать о том, что для ценопопуляции в данных условиях зафиксированные максимальные значения не являются аномально высокими.
У эпилитного лишайника Protoparmeliopsis muralis в размерном ряду талломов от 1 до 26 см2 за 11-летний период наблюдения выявлен только линейный рост. Ряд авторов вслед за М. Бешелем (Beschel, 1960) выделяют долинейную фазу роста, в которой у талломов постепенно увеличиваются годовые приросты, и постлинейную, в которой по мере старения таллома годовые приросты снижаются (Platt, Amsler, 1955; Beschel, 1960; Armstrong, 1974).
Существуют данные, что долинейная фаза роста может протекать наиболее быстро по сравнению с другими фазами роста, к примеру, у лишайников рода Rizocarpon она завершается, когда талломы еще имеют микроскопические размеры (Innes, 1985). Линейная и постлинейная фазы роста сильно отличаются по продолжительности в зависимости от биологических особенностей вида лишайника и условий окружающей среды. По мере старения лишайника скорость его роста замедляется, может снижаться до минимальных значений или останавливаться (Галанин, Глушкова, 2003). По достижении максимальных размеров, характерных для особи, отдельные участки талломов начинают разрушаться. Разрушения талломов также происходят вследствие влияния популяционно-ценотических и сукцессионных факторов (например, при контакте с более конкурентоспособными видами), выветривании и разрушении поверхности субстрата (Галанин, 2001).
Долинейная фаза роста может происходить у талломов Protoparmeliopsis muralis на начальных этапах онтогенеза, когда размеры таллома менее 1 см2. Учитывая сравнительно небольшую продолжительность жизни данного вида, можно предположить, что остановка роста талломов может происходить достаточно быстро, как и полное разрушение талломов, некоторые талломы полностью разрушались в течение года между наблюдениями.
Растущие талломы Protoparmeliopsis muralis могут частично разрушаться, подвергаясь воздействию как абиотических факторов (например, образование ледяной корки, сход снега и льда на реке), так и биотических в силу конкурентной борьбы с другими видами лишайников. Чаще всего разрушению подвержены краевые части талломов, имеющие лопастное строение. Лопастные участки талломов неплотно связаны с субстратом и более подвержены разрушению, чем центральная чешуйчатая часть. В дальнейшем разрушенные участки могут восстанавливаться. У талломов, близких к своим максимальным размерам и возрасту, кроме разрушения краевых частей часто отмечалось начало разрушения таллома в центральной части.
Учитывая проведенный анализ роста всех талломов вида Protoparmeliopsis muralis в диапазоне от 1 до 26 см2 в прибрежном эпилитном сообществе на отдельном фрагменте скалы, можно предположить, что большую часть жизни талломы растут линейно. Реальный возраст талломов несколько больше оцененной продолжительности линейного роста на то время, которое необхоям2, и время от остановки роста талломов до полного его разрушения. Предположительно продолжительность начального роста и время до полного разрушения талломов должны быть меньше зафиксированного линейного роста талломов.
Полученные данные свидетельствуют о небольшой продолжительности жизни вида в условиях Карелии и невозможности использования данного вида в лихенометрическом датировании.
Заключение
В настоящем исследовании впервые получены данные о росте эпилитного лишайника Protoparmeliopsis muralis, обитающего на приречных скалах в условиях таежной зоны Карелии. На протяжении исследования размеры талломов находились в диапазоне 1…26 см2. В пределах зафиксированных размеров талломов отмечен только линейный рост. Модельные скорости роста талломов варьируют от 0.2 до 1.50 см2. Долинейная и постлинейная фазы роста нами не были выявлены. Оцененная по моделям продолжительность линейного роста составляет 6…23 года. Реальный возраст талломов несколько больше полученной продолжительности линейного роста на неучтенное в моделях время, необходимое для развития талломов от микроскопических размеров до 1 см2, и время от остановки роста до разрушения талломов. В связи с небольшой продолжительностью жизни на прибрежных скалах в эпилитных синузиях вид непригоден для лихеонометрического датирования скальных субстратов в данном регионе.
Библиография
Викторов С. В. Лишайники как индикаторы литологических и геохимических условий в пустыне // Вестник Московского университета. Сер.: Биология. 1956. № 5. C. 115–119.
Викторов С. В. Лишайники пустыни Устюрт и их связь с некоторыми свойствами почв и горных пород // Вопросы индикационной геоботаники. М., 1960. C. 53–60.
Галанин А. А. Лихенометрические кривые роста Rhizocarpon sp. в горах Северо-Востока Азии и Северной Америки // Проблемы геологии и металлогении Северо-Востока Азии на рубеже тысячелетий: четвертичная геология, геоморфология, россыпи. Магадан: СВКНИИ ДВО РАН, 2001. С. 11–14.
Галанин А. А. Лихенометрический метод в изучении современных геоморфологических процессов на Северо-Востоке России : Автореф. дис. … канд. геогр. наук. Владивосток: ТИГ ДВО РАН, 1997. 21 с.
Галанин А. А. Лихенометрический метод изучения криогенных процессов // Наука и техника в Якутии. 2012. № 1. С. 8–15.
Галанин А. А., Глушкова О. Ю. Лихенометрия // Вестник РФФИ. 2003. № 3. С. 3–38.
Галанина И. А. К вопросу о скорости роста лишайников-эпилитов на юге Приморского края // Структура и динамика экосистем Сибири и Дальнего Востока: Сб. науч. ст. Находка: Институт технологии и бизнеса, 2011. С. 130–143.
Домбровская А. В. Влияние некоторых экологических факторов на распределение и рост кустистых и листоватых лишайников в Хибинах // Ботанический журнал. 1963. Т. 48. № 5. С. 742–748.
Михайлова И. Н. Анализ субпопуляционных структур эпифитных лишайников (на примере Lobaria pulmonaria (L.)) // Вестник Нижегородского университета им. Н. И. Лобачевского. Сер.: Биология. 2005. № 1. С. 124–134.
Ивантер Э. В., Коросов А. В. Введение в количественную биологию . Петрозаводск, 2003. 304 с.
Турманина В. И. Пути восстановления природных условий последнего тысячелетия и основные результаты // Ритмы природных процессов. М.: МГУ, 1979. С. 24–55.
Фадеева М. А., Сонина А. В. Лихенологические исследования онежских петроглифов в связи с оценкой их состояния // Труды Первой российской лихенологической школы. 2000. С. 238–250.
Фадеева М. А., Сонина А. В. Скорость роста эпилитных лишайников как показатель деструктивного воздействия лихенобиоты на онежские петроглифы // Актуальные проблемы геоботаники: современные направления исследований в России: методологии, методы и способы обработки материалов: Тез. докл. Петрозаводск, 2001. С. 188–189.
Andrews J. T., Webber P. J. A lichenometrical study of the northwestern margin of the Barnes Ice Cap // A geomorphological technique. Geogr. Bull. 1964. № 22. P. 80–104.
Armstrong R. A. Growth phases in the life of a lichen thallus // New Phytologist. 1974. № 73. P. 913–918.
Beschel R. E. A project to use Lichens as indicators of climate and time // Arctic Alpine Res. 1957. Vol. 10. № 1. P. 200–215.
Beschel R. E. Dating rock surfaces by growth and its application to glaciology and physiography (lichenometry) // Geology of the Arctic: proceedings of the first international symposium on Arctic Geology. 1960. P. 1044–1062.
Calkin P. E., Ellis J. M. A lichenometric dating curve and its application to Holocene glacier studies in the central Brooks Range, Alaska // Arctic and Alpine Research. 1980. Vol. 12. № 3. P. 245–264.
Denton G. H., Karlén W. Lichenometry: its application to Holocene moraine studies in southern Alaska and Swedish Lapland // Arctic and Alpine Research. 1973. Vol. 5. P. 347–372.
Farrar J. F. A method for investigating lichen growth rates and succession // The Lichenologist. 1974. Vol. 6. № 2. С. 151–155.
Innes J. L. Lichenometry // Progress in physical geography. 1985. Vol. 9. № 2. P. 187–254.
Miller G. H., Andrews J. T. Quaternary history of northern Cumberland Peninsula, East Baffin Island, NWT, Canada Part VI: preliminary lichen growth curve for Rhizocarpon geographicum // Geological Society of America Bulletin. 1972. Vol. 83. № 4. P. 1133–1138.
Platt R. B., Amsler F. P. A basic method for the immediate study of lichen growth rates and succession // J. Tenn. Acad. Sci. 1955. Vol. 30. P. 177–183.
Purvis O. W., Coppins B. J., Hawksworth D. L., James P. W., More D. M. Lichen Flora of Great Britain and Ireland. Natural History Museum Publications in association with the British Lichen Society, 1992. 710 p.
Reger R. D., Pewe T. L. Lichenometric dating in the central Alaska Range // The Periglacial Environment. 1969. P. 223–247.

 © 2011 - 2026
© 2011 - 2026