



Введение
Строение жевательной поверхности коренных зубов полевок традиционно используется в таксономических (Огнев, 1940; Громов, Ербаева, 1995) и микроэволюционных исследованиях (Большаков и др., 1980; Chaline et al., 1993), поскольку для строения коренных зубов характерна, с одной стороны, видоспецифичность, с другой – значительная изменчивость (Runck et al., 2009; Дубинин, 2016).
В настоящее время существует несколько подходов к оценке сложности жевательной поверхности полевок подсемейства Arvicolinae, учитывающие такие признаки, как число, форма, положение и степень слияния элементов жевательной поверхности (Маркова, 2013). Наиболее удобным, на наш взгляд, является морфотипический анализ с выделением вариаций строения жевательной поверхности – морфотипов – и оценкой их встречаемости в популяциях. Схемы морфотипической изменчивости разработаны для большинства современных видов (Ангерманн, 1973; Большаков и др., 1980; Поздняков, 1993 и др.) и ископаемых форм полевок (Малеева, 1976; Малеева, Шувалова, 1980; Смирнов и др., 1986). Применение морфотипического анализа позволяет использовать результаты изучения современных животных в качестве сравнительной базы для исследования ископаемых остатков, а также для выяснения вопросов о происхождении и эволюции видов (Малеева, 1976).
Морфотипическая изменчивость жевательной поверхности коренных зубов полевок – это случай морфологического полиморфизма, при котором наблюдается направленный сдвиг во времени и пространстве преобладающего типа строения, в данном случае – строения жевательной поверхности коренных зубов. Она отражает генетическое многообразие вида, накопившееся за всю историю его развития. По различным проявлениям морфотипической изменчивости можно понять, как вид приспособился к условиям существования, и оценить характер давления естественного отбора в процессе формирования вида (Большаков и др., 1980). Подобную изменчивость удобно описывать в виде ряда морфотипов (Ларина, Еремина, 1988), а расчет частот встречаемости морфотипов дает возможность говорить о морфотипической изменчивости популяции и позволяет получить представление о вызывающих ее факторах. Анализ ряда работ по морфотипической изменчивости зубов корнезубых полевок показывает, что данная морфологическая характеристика находится под влиянием нескольких факторов. По мнению Т. А. Андреевой (2008), изменчивость структуры жевательной поверхности М3 связана с географическими особенностями территорий в пределах ареалов видов: например, для M. rutilus установлено усложнение строения коренных зубов в восточном направлении, а для M. glareolus – в северном. А. В. Бобрецов (2010) отмечает, что на территории Печеро-Илычского заповедника частотный состав морфотипов меняется в ландшафтном градиенте, а именно: частота сложных морфотипов М3 увеличивается от равнины к горам. Рядом авторов отмечается связь между структурой жевательной поверхности М3 и питанием на основании анализа содержимого желудков (соотношение зеленых и семенных кормов) M. glareolus с рисунком жевательной поверхности зуба, при этом подчеркивается адаптивная значимость этих структур (Воронцов, 1967; Емельянова, 2008; Окулова, Андреева, 2008). При увеличении доли зеленых кормов в рационе M. glareolus показано увеличение частоты встречаемости простых морфотипов жевательной поверхности коренных зубов. Кроме этого изменчивость структуры жевательной поверхности зубов подвержена возрастной изменчивости (Коурова, 1986; Бородин и др., 2006; Емельянова, 2008). Таким образом, данные факторы оказывают существенное влияние на морфотипическую изменчивость зубов полевок.
Учитывая широкое географическое распространение и высокое обилие в бореальных экосистемах Тюменской области, красная полевка (Myodes rutilus Pallas, 1779) представляет собой удобную модель для изучения морфотипической изменчивости.
В настоящее время региональные закономерности проявления морфотипической изменчивости даже у широко распространенных видов остаются малоизученными, поэтому цель нашей работы – описание морфотипической изменчивости жевательной поверхности зуба М3 Myodes rutilus на территории Среднего Приобья Тюменской области, изучение возрастной изменчивости и изменчивости структуры жевательной поверхности зуба М3, связанной с фазами популяционного цикла и биотопическими особенностями местообитания зверьков.
Материалы
Материалом для нашей работы послужили коллекционные сборы черепов M. rutilus на территории Среднего Приобья Тюменской области с 1987 по 1990 г., хранящихся в Зоологическом музее ТюмГУ. Схема сбора материала представлена на рис. 1, распределение объема выборки по точкам сбора и частоты морфотипов М3 – в табл. 1. В ходе работы всего было просмотрено 538 черепов красной полевки. Для исследования морфотипической изменчивости структуры жевательной поверхности был использован третий верхний коренной зуб М3. В основу классификации морфотипов жевательной поверхности зуба М3 была положена методика И. В. Ереминой (1981) с дополнениями (Окулова, Андреева, 2008), разработанная для рыжей полевки. На основании близкого родства красной и рыжей полевок мы сочли возможным использование данных методик и для красной полевки.
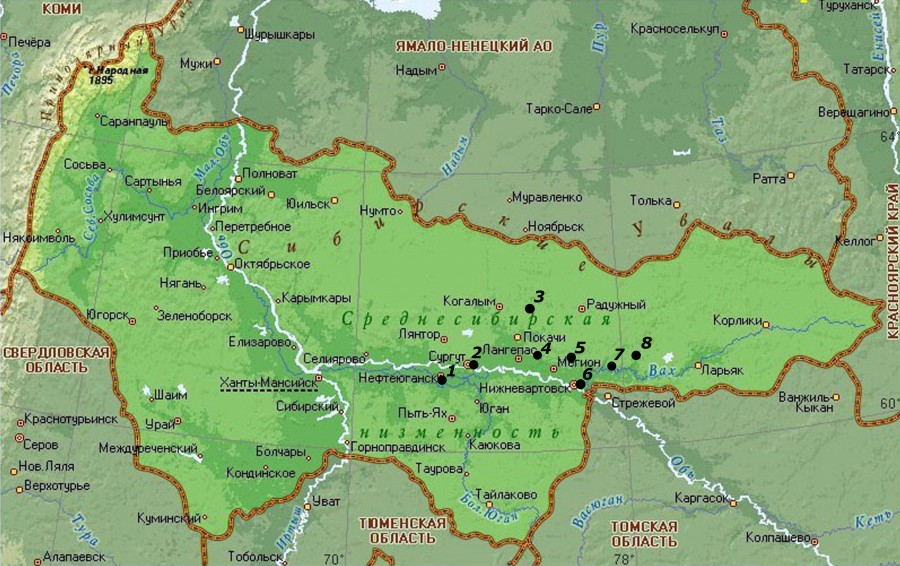
Рис. 1. Схема сбора материала:
1 – г. Нефтеюганск (Нефтеюганский р-н), 2 – г. Сургут (Сургутский р-н), 3 – п. Аган (Сургутский р-н), 4 – п. Ватинск (Нижневартовский р-н), 5 – Мыхпайское месторождение (Нижневартовский р-н), 6 – г. Нижневартовск (Нижневартовский р-н), 7 – р. Пасол (Нижневартовский р-н), 8 – р. Вах (Нижневартовский р-н)
Fig. 1. The scheme of collecting material:
1 – Nefteyugansk (Nefteyugansky district), 2 – Surgut (Surgut district), 3 – Agan village (Surgut district), 4 – Vatinsk (Nizhnevartovsk district), 5 – Myhpayskoye field (Nizhnevartovsk district), 6 – Nizhnevartovsk (Nizhnevartovsk district), 7 – r. Pasol (Nizhnevartovsk district), 8 – r. Vakh (Nizhnevartovsk district)
Таблица 1. Частоты морфотипов М3 красной полевки
| № | Место сбора материала | n | Число выбо-рок | Частоты морфотипов М3 | ||||||
| a | b | c | e | f | glf | ss | ||||
| 1 | г. Нефтеюганск (Нефтеюганский р-н) | 11 | 1 | 0.091 | - | 0.818 | 0.091 | - | - | |
| 2 | г. Сургут (Сургутский р-н) | 18 | 1 | 0.056 | - | 0.778 | 0.167 | - | - | - |
| 3 | п. Аган (Сургутский р-н) | 230 | 14 | 0.018 | 0.014 | 0.764 | 0.193 | 0.004 | 0.002 | 0.006 |
| 4 | п. Ватинск (Нижневартовский р-н) | 47 | 5 | 0.101 | - | 0.712 | 0.187 | - | - | - |
| 5 | Мыхпайское месторождение (Нижневартовский р-н) | 20 | 2 | 0.056 | - | 0.889 | 0.056 | - | - | - |
| 6 | г. Нижневартовск (Нижневартовский р-н) | 84 | 3 | 0.017 | 0.006 | 0.895 | 0.071 | 0.006 | 0.006 | - |
| 7 | р. Пасол (Нижневартовский р-н) | 37 | 2 | 0.180 | - | 0.578 | 0.124 | - | 0.019 | 0.100 |
| 8 | р. Вах (Нижневартовский р-н) | 91 | 7 | 0.022 | - | 0.712 | 0.266 | - | - | - |
Примечание. n – объем выборки, a – ss – частота встречаемости морфотипов.
Методы
Строение жевательной поверхности зуба М3 (левый зубной ряд) рассматривали с помощью бинокулярного микроскопа МБС при увеличении в 20 раз. Анализ морфотипической изменчивости проводился для неполовозрелых, достигших размеров взрослых, и взрослых половозрелых особей, как самцов, так и самок, для которых характерны зубы с полностью раскрытой от эмали поверхностью. Старые и ювенильные особи использовались только при оценке разнообразия морфотипов и изучении возрастной изменчивости. Для изучения возрастной изменчивости все зверьки были разделены на две возрастные группы: сеголетки и зимовавшие. Возраст зверьков определялся по степени редукции альвеолярного бугра (Кошкина, 1955). При выделении дискретных вариантов (морфотипов) зуба М3 пользовались методикой Окуловой, Андреевой (2008), где учитывалось количество входящих и выходящих углов на лингвальной и буккальной сторонах зуба М3 (рис. 2). После были рассчитаны частоты встречаемости морфотипов в выборке и коэффициенты сложности зуба M3 (КСЗ). Подсчет коэффициентов сложности зуба вели, придав баллы сложности морфотипам зуба M3: a – 1, b – 2, c – 3, d – 4, e – 5, f – 6, более сложным – 7. Затем рассчитывали КСЗ по следующей формуле: КСЗ = Ba × Fa + Bb × Fb + Bc × Fc + Bd × Fd + Be × Fe и т. д., где В – баллы сложности данного морфотипа, а F – частота встречаемости данного морфотипа. Статистическая обработка данных проводилась с применением программного пакета StatSoft Statistica 10.
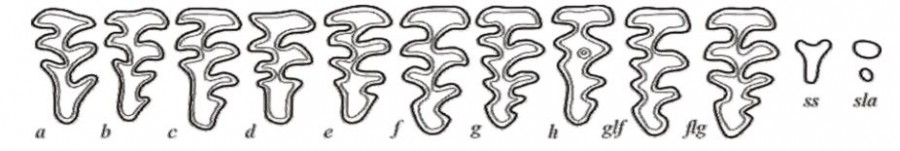
Рис. 2. Вариации рисунка жевательной поверхности зуба М3
Fig. 2. Variations of the chewing surface pattern of the М3 tooth
Результаты
В популяции красной полевки, обитающей на территории Среднего Приобья Тюменской области, были обнаружены 7 (a, b, c, e, f, glf, ss) из 12 известных морфотипов зуба М3, что свидетельствует о невысоком разнообразии морфотипов.
Морфотип а встретился у 4.46 % особей (на М3 имеется два входящих угла на лингвальной стороне зуба) и только у единичных особей. Как правило, у данного вида полевок преобладают морфотипы, зуб М3 которых характеризуется наличием трех входящих углов на лингвальной стороне (морфотипы b, c, e, f). Этот признак является диагностическим (Громов, Ербаева, 1995; Павлинов и др., 2002) и он присутствует у 94.42 % полевок. Однако только у 71.75 % особей имеется типичное для красной полевки строение М3, а именно: наличие двух входящих углов на буккальной стороне и трех входящих углов на лингвальной стороне зуба (морфотипы b и c). У 5.02 % особей зуб М3 имеет иное количество входящих углов. Морфотипы e, f имеются у 22.67 % особей красной полевки, для них характерны по три входящих угла с обеих сторон зуба. Морфотип glf имеет четыре входящих угла на наружной стороне и три на внутренней стороне М3 и был обнаружен лишь у 0.56 % особей. Морфотип ss характеризуется редукцией входящих и исходящих углов вследствие истирания и встречается также у 0.56 % особей (табл. 2).
Таблица 2. Разнообразие морфотипов зуба М3 M. rutilus Среднего Приобья
| n | Морфотипы жевательной поверхности зуба М3 | КСЗ | |||||||
| a | b | c | e | f | glf | ss | |||
| Количество особей | 538 | 24 | 8 | 378 | 118 | 4 | 3 | 3 | |
| Процентное соотношение, % | 4.46 | 1.49 | 70.26 | 21.93 | 0.74 | 0.56 | 0.56 | 3.40 | |
Примечание: n – количествово особей.
В соответствии с методикой, все морфотипы зуба М3 были распределены следующим образом: к группе «простых» отнесены морфотипы a и b, к группе «средней сложности» – c и d (в нашем случае морфотип d отсутствует), к группе «сложные» – e, f, glf и ss.
Морфотипическая изменчивость жевательной поверхности зуба М3 характеризуется невысокой частотой встречаемости простых морфотипов (5.95 %). Наиболее часто встречающимися являются морфотипы средней сложности, частота встречаемости которых равна 70.26 %, частота встречаемости сложных морфотипов –23.79 % (табл. 3).
Таблица 3. Частоты морфотипов зуба М3 M. rutilus Среднего Приобья
| n | Морфотипы жевательной поверхности зуба М3 | |||
| простые a + b | средняя степень сложности c | сложные e + f и сложнее |
||
| Количество особей | 538 | 32 | 378 | 128 |
| Процентное соотношение морфотипов М3, % | 100 | 5.95 | 70.26 | 23.79 |
Примечание. n – объем выборки.
Для изучения возрастной изменчивости зуба М3 было проведено сравнение частот встречаемости морфотипов разной степени сложности и КСЗ в группах зимовавших особей и сеголеток красной полевки. Анализ особенностей строения М3 показал, что у зимовавших особей частота встречаемости простых морфотипов и морфотипов средней сложности достоверно выше, чем у сеголеток. В группе зимовавших особей отмечается достоверное увеличение доли морфотипов средней сложности при достоверном снижении доли сложных морфотипов, в связи с чем КСЗ М3 зимовавших особей красной полевки значительно ниже, чем у сеголеток (табл. 4).
Таблица 4. Средние частоты морфотипов зуба М3 (M ± m) M. rutilus Среднего Приобья в различных возрастных группах
| Возрастная группа | n | Частоты морфотипов М3 | КСЗ | ||
| простые a + b | средняя степень сложности c | сложные e + f и сложнее |
|||
| Зимовавшие | 121 | 0.102 ± 0.027* | 0.831 ± 0.041*** | 0.067 ± 0.048*** | 2.81 |
| Сеголетки | 405 | 0.049 ± 0.026 | 0.675 ± 0.038 | 0.277 ± 0.025 | 3.53 |
Примечание. n – объем выборки, * – различия достоверны при р < 0.05, *** – различия достоверны при р < 0.001.
В период сбора материала, с 1987 по 1990 г., была выявлена динамика относительного обилия красной полевки. Данный показатель достигает высоких значений (14.03 экз./100 лов.-сут.) в 1987 г., в последующие 2 года отмечено последовательное снижение относительного обилия зверьков, а в 1989 г. оно было минимальным – 2.93 экз./лов.-сут., что, вероятно, связано с аномально высокими летними температурами. В 1990 г. относительное обилие вида значительно увеличивается до 14.93 экз./лов.-сут.
При изучении разнообразия морфотипов зуба М3 красной полевки, в зависимости от показателя относительного обилия зверьков, было выявлено, что морфотипы a, c и e встречались в течение всего популяционного цикла 1987–1990 гг., следовательно, они являются основой морфотипического разнообразия жевательной поверхности М3 красной полевки в исследуемый период (табл. 5). Морфотип f встречается единично практически во все годы, за исключением 1989 г., с наименьшим относительным обилием. Только в 1990 г. отсутствует морфотип b и присутствует морфотип glf. Таким образом, в год, характеризующийся депрессией численности красной полевки (1989), отмечено уменьшение разнообразия морфотипов, однако это может быть связано со снижением объема выборки.
Таблица 5. Разнообразие морфотипов зуба М3 M. rutilus Среднего Приобья в период 1987–1990 гг.
| Год | N | n | Морфотипы М3 | |||||
| a | b | c | e | f | glf | |||
| 1987 | 14.03 | 140 | + | + | + | + | + | - |
| 1988 | 4.33 | 131 | + | + | + | + | + | - |
| 1989 | 2.93 | 64 | + | + | + | + | - | - |
| 1990 | 14.93 | 146 | + | - | + | + | + | + |
Примечание. N – относительное обилие красной полевки (экз./100 лов.-сут.), n – количество особей, + – присутствие морфотипа в выборке, - – отсутствие морфотипа в выборке.
Для красной полевки Среднего Приобья выявлено изменение частот встречаемости морфотипов зуба М3 в ходе популяционных циклов, характеризующихся различным относительным обилием (табл. 6). Частоты встречаемости морфотипов средней сложности (c) на всех фазах популяционного цикла с 1987 по 1990 г. были относительно стабильны. Снижение относительного обилия красной полевки (1988 и 1989 гг.) сопровождалось достоверным увеличением частоты встречаемости простых морфотипов (a + b) зуба М3. А в годы с высоким относительным обилием красной полевки (1987 и 1990) отмечается достоверно низкая частота встречаемости простых морфотипов.
Таблица 6. Средние частоты морфотипов зуба М3 (M ± m) M. rutilus Среднего Приобья в период 1987–1990 гг.
| Год | N | Частоты морфотипов М3 | |||
| простые a + b | средняя степень сложности c | сложные e + f и сложнее |
КСЗ | ||
| 1987 | 14.03 | 0.065 ± 0.034 * | 0.722 ± 0.011 | 0.212 ± 0.046 ● | 3.41 |
| 1988 | 4.33 | 0.082 ± 0.017 ♦♦♦ | 0.711 ± 0.042 | 0.207 ± 0.032 | 3.31 |
| 1989 | 2.93 | 0.095 ± 0.006●● | 0.734 ± 0.112 | 0.171 ± 0.002 | 3.19 |
| 1990 | 14.93 | 0.040 ± 0.027 | 0.789 ± 0.020 | 0.170 ± 0.040 | 3.40 |
Примечание. N – относительное обилие красной полевки (экз./100 лов.-сут.), * – сравнение 1987 и 1988 гг., ♦ – сравнение 1988 и 1990 гг., ● – сравнение 1989 и 1990 гг., различия достоверны при: 1 знак – р < 0.05, 2 знака – р < 0.01, 3 знака – р < 0.001.
Также была выявлена связь между показателем относительного обилия красной полевки и коэффициентом сложности зуба М3 (рис. 3). Изменения частот встречаемости зубов с различной сложностью находят отражение в значениях КСЗ М3 (см. табл. 6). Поскольку большинство выборок отклонялись от нормального распределения, нами был использован непараметрический метод – расчет коэффициента корреляции Спирмена (rsp). В результате проведенного анализа была выявлена достоверная положительная связь между величиной относительного обилия и КСЗ М3: коэффициент корреляции Спирмена - rsp = 0.79 ± 0.02 при р < 0.05.
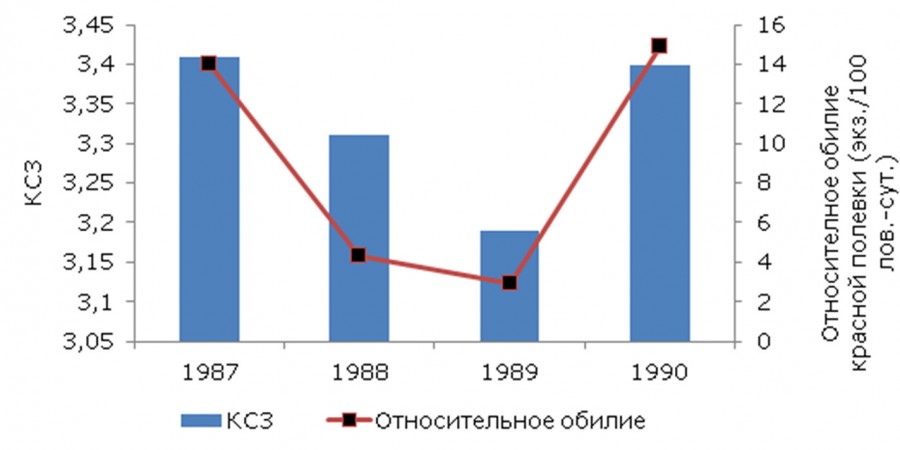
Рис. 3. Зависимость КСЗ М3 от относительного обилия красной полевки Среднего Приобья
Fig. 3. The dependence of the complexity factor of the M3 tooth on the relative abundance of the red vole in the Middle Ob area
Красная полевка широко распространена в средней тайге и способна заселять самые различные по степени благоприятности местообитания. Для анализа влияния биотопического распределения зверьков на морфотипы зуба М3 красной полевки были взяты типичные для средней тайги биотопы: разнотравные ассоциации, осоковые болота, производные леса (опушки, гари, поросли), долгомошные ассоциации, приручьевые и травяно-болотные ассоциации.
Анализ данных табл. 7 показал изменение частот встречаемости морфотипов различной сложности зуба и КСЗ М3 красной полевки для местообитаний с различными биотопическими особенностями. Наиболее высокий коэффициент сложности зуба М3 наблюдается у полевок, обитающих в разнотравных ассоциациях (3.54) и в осоковых болотах (3.48), где усложнение зуба происходит за счет высокой частоты встречаемости сложных морфотипов (0.318). Зеленомошные, травяно-болотные и приручейниковые ассоциации характеризуются наибольшей частотой встречаемости простых морфотипов и невысокими частотами сложных морфотипов, вследствие чего КСЗ М3 имеет низкие значения. Производные леса и долгомошные ассоциации занимают промежуточное положение по значению КСЗ М3.
Таблица 7. Средние частоты морфотипов зуба М3 (M ± m) красной полевки
Среднего Приобья в различных местообитаниях
| Биотоп | n | Частоты морфотипов М3 | КСЗ | ||
| простые a + b | средняя степень сложности c | сложные e + f и сложнее |
|||
| Разнотравные ассоциации | 48 | 0.121 ± 0.091 | 0.652 ± 0.035 | 0.318 ± 0.041 | 3.54 |
| Осоковые болота | 21 | 0 | 0.743 ± 0.019 | 0.257 ± 0.019 | 3.48 |
| Производные леса | 43 | 0.036 ± 0.021 | 0.839 ± 0.059 | 0.125 ± 0.072 | 3.35 |
| Долгомошные ассоциации | 93 | 0.114 ± 0.012 | 0.676 ± 0.068 | 0.211 ± 0.017 | 3.33 |
| Приручейниковые ассоциации | 38 | 0.054 ± 0.004 | 0.720 ± 0.041 | 0.225 ± 0.043 | 3.32 |
| Травяно-болотные ассоциации | 84 | 0.082 ± 0.005 | 0.727 ± 0.003 | 0.191 ± 0.001 | 3.30 |
| Зеленомошные ассоциации | 108 | 0.038 ± 0.024 | 0.827 ± 0.048 | 0.135 ± 0.039 | 3.26 |
Примечание. n – кол-во особей.
Обсуждение
На основании долевого участия отдельных морфотипов был рассчитан коэффициент сложности зуба (КСЗ) М3. КСЗ М3 красной полевки, обитающей в подзоне средней тайги Среднего Приобья Тюменской области, равен 3.40 (см. табл. 1). По результатам наших исследований (Сорокина, 2011), для популяции красной полевки в северной тайге Тюменской области значения КСЗ М3 выше и составляют 3.66. По данным Сорокиной и Сидоровой (2016), величина КСЗ М3 красной полевки в северной тайге Среднего Зауралья составила 3.76, а в подтайге – 3.09. Коэффициент сложности зуба М3 красной полевки в южной тайге Европейской части России (Удмуртия) равен 2.88 (Окулова, Андреева, 2008). Таким образом, при сравнении коэффициентов сложности зуба М3 красной полевки в вышеуказанных подзонах можно отметить, что КСЗ М3 красной полевки средней тайги в исследуемом районе хорошо вписывается в ряд, характеризующий географическую изменчивость строения жевательной поверхности М3 и отражает последовательное усложнение жевательной поверхности этого зуба в северном направлении.
Об изменении морфотипических характеристик зубов полевок на разных возрастных стадиях неоднократно упоминалось в литературе (Воронцов, 1967; Андреева, Окулова, 2005; Емельянова, 2005; Бородин и др., 2006). Для красной полевки, обитающей в подзоне средней тайги Среднего Приобья, также характерна возрастная изменчивость структуры жевательной поверхности зуба М3. Упрощение строения зуба М3 красной полевки по мере увеличения возраста, вероятно, связано со стачиванием призмы зуба в течение жизни. Поскольку входящие углы моляров залегают на неодинаковые глубины по всей высоте коронки и в прикорневой части они наименьшие, следовательно, при стирании коронки уменьшается количество входящих углов, а значит, упрощается строение зуба. Следовательно, строение коренных зубов М3 красной полевки подвержено возрастной изменчивости.
Изучение частот встречаемости различных по сложности морфтотипов в ходе популяционных циклов показало, что морфотип средней сложности (с) характеризуется относительно постоянной частотой встречаемости, поэтому можно полагать, что они обладают наибольшей адаптивной ценностью. Увеличение частот встречаемости простых морфотипов (a + b) жевательной поверхности зуба М3 связано с комплексом адаптаций, повышающих выживаемость зверьков в условиях низкого относительного обилия. Возможно, простые морфотипы имеют адаптивное значение для популяции красной полевки в период с низким относительным обилием вида при стабильной частоте встречаемости морфотипов средней сложности. Изменение частоты встречаемости морфотипов на разных фазах динамики численности отмечено и для рыжей полевки Т. А. Андреевой (2008).
Выявленная достоверная положительная связь между показателем относительного обилия красной полевки и коэффициентом сложности зуба М3 свидетельствует о том, что при снижении относительного обилия красной полевки отмечается более простое строение жевательной поверхности зуба М3, и наоборот: с увеличением относительного обилия вида наблюдается наиболее сложное строение жевательной поверхности зуба М3. Аналогичная закономерность прослеживается в работах Т. А. Андреевой (2008), которая изучала изменчивость коренного зуба М3 рыжей полевки в различных частях ареала вида. Возможно, выявленная тенденция зависимости КСЗ зуба М3 от относительного обилия красной полевки связана с переживанием неблагоприятных условий (в первую очередь климатических). Т. А. Андреева, Н. М. Окулова (2005) отмечают сходную тенденцию: увеличение частоты встречаемости простых морфотипов М3 рыжей полевки Приокско-Террасного заповедника в засушливое лето.
Исследование изменения частот встречаемости морфотипов различной сложности зуба и КСЗ М3 красной полевки для местообитаний с различными биотопическими особенностями показало определенную зависимость. Анализ литературных данных свидетельствует о том, что наиболее богатыми по числу видов и относительному обилию мелких млекопитающих являются зеленомошные, травяно-болотные и приручейниковые ассоциации (Гашев, 1991). По нашим данным, в вышеперечисленных местообитаниях наблюдается невысокая сложность жевательной поверхности зуба М3 красной полевки. Кроме этого, С. Н. Гашев (1996) утверждает, что зеленомошные кедровники являются для красной полевки наиболее благоприятными местообитаниями, а также в зеленомошниках красная полевка переживает неблагоприятные годы и (или) сезоны, и относительное обилие вида здесь меняется незначительно, даже в годы депрессий. Работы Э. В. Ивантера и Е. А. Моисеевой (2015) подтверждают предположение о том, что красная полевка в условиях Карелии также отдает предпочтение хвойным зеленомошным лесам.
Следовательно, степень благоприятности местообитаний оказывает влияние на сложность строения жевательной поверхности зуба М3, т. е. в наиболее благоприятных условиях структура жевательной поверхности зуба М3 имеет меньшее число входящих углов, в менее благоприятных – большее, что ведет к усложнению строения зуба. Данное предположение основано на уже изученной зависимости структуры жевательной поверхности от биотопических особенностей и состава растительного покрова и, следовательно, от рациона питания зверьков (Емельянова, Суворова, 2008; Souto-Lima, Millien, 2014; Ямборко, 2015).
Заключение
При исследовании жевательной поверхности М. rutilus Среднего Приобья Тюменской области были выявлены следующие морфотипы: a, b, c, e, f, glf, ss. 94.42 % встреченных морфотипов характеризуются тремя входящими углами на лингвальной стороне зуба М3; у 5.58 % особей количество входящих углов на лингвальной стороне иное (либо два, либо четыре).
Для М. rutilus Среднего Приобья выявлена возрастная изменчивость строения жевательной поверхности зуба М3. У зимовавших особей жевательная поверхность зуба М3 имеет более простую структуру по сравнению с сеголетками вследствие истирания коронки зуба.
Морфотипическая изменчивость структуры жевательной поверхности М3 красной полевки проявляется в достаточно стабильной частоте встречаемости морфотипов средней сложности при повышении частоты встречаемости простых морфотипов в годы с низким относительным обилием вида. Изменения частотного состава морфотипов находят отражение и в КСЗ М3.
На строение жевательной поверхности М3 красной полевки Среднего Приобья влияют биотопические особенности местообитаний. В наиболее благоприятных условиях структура жевательной поверхности характеризуется меньшей сложностью, в менее благоприятных отмечена тенденция усложнения строения зуба.
Библиография
Ангерманн Р. Гомологическая изменчивость коренных зубов у полевок (Microtinae) // Проблемы эволюции. Новосибирск: Наука, 1973. Т. З. С. 104–119.
Андреева Т. А., Окулова Н. М. Изменчивость рыжей полевки Приокско-Террасного заповедника // Приокско-Террасный государственный природный биосферный заповедник. Экосистемы Приокско-Террасного биосферного заповедника. Пущино, 2005. С. 160–167.
Андреева Т. А. Внутривидовая дифференциация европейской рыжей полевки Clethrionomys glareolus Sshr., 1780 : Автореф. дис. … канд. биол. наук. М.: Московский гос. университет им. М. В. Ломоносова, 2008. 26 с.
Бобрецов А. В. Морфотипическая изменчивость зубов красной полевки (Clethrionomys rutilus) Печоро-Илычского заповедника // Труды Печоро-Илычского заповедника. 2010. С. 10–17.
Большаков В. Н., Васильева И. А., Малеева А. Г. Морфотипическая изменчивость зубов полевок . М.: Наука, 1980. 140 с.
Бородин А. В., Давыдова Ю. А., Елькина М. А. Одонтологические характеристики полевок рода Clethrionomys (Tilesius, 1850) Висимского заповедника // Экологические исследования в Висимском биосферном заповеднике: Материалы научной конференции, посвященной 35-летию Висимского заповедника. Екатеринбург, 2–3.10.2006 г. Екатеринбург, 2006. С. 73–81.
Воронцов Н. Н. Эволюция пищеварительной системы грызунов (Мышеобразные) . Новосибирск, 1967. 240 с.
Гашев С. Н. Влияние нефтяного загрязнения на фауну и экологию мелких млекопитающих Среднего Приобья : Автореф. дис. … канд. биол. наук. Екатеринбург: Институт экологии растений и животных УрО АН СССР, 1991. 25 с.
Гашев С. Н. Состояние фауны мелких млекопитающих Среднего Приобья // Биоразнообразие Западной Сибири – результаты исследований / Под ред. В. Р. Цибульского. Тюмень: Изд-во Института проблем освоения Севера СО РАН, 1996. С. 9–16.
Громов И. М., Ербаева М. А. Млекопитающие фауны России и сопредельных территорий: Зайцеобразные и грызуны . СПб.: Зоологический институт РАН, 1995. 522 с.
Дубинин Е. А. Темпоральная изменчивость строения жевательной поверхности М3 красной полевки северо-востока Сибири // Вестник Северо-восточного государственного университета. 2016. Вып. 25. С. 38–46.
Емельянова А. А. Возрастная изменчивость одонтологических признаков рыжей полевки (Clethryonomys glareolus Schreber) // Вестник Тверского государственного университета. Серия: Биология и экология. 2005. Вып. 1. № 4 (10). С. 88–96.
Емельянова А. А. Питание европейской рыжей полевки верховий Волги и смежных территорий // Вестник ТвГУ. 2008. № 10. С. 109–118.
Емельянова А. А., Суворова А. А. Некоторые закономерности полиморфической изменчивости одонтологических признаков европейской рыжей полевки (Clethrionomys glareolus), обитающей в верховьях Волги и на сопредельных территориях // Вестник ТвГУ. 2008. № 7. С. 79–88.
Еремина И. В. Вариации строения моляров рецентных и ископаемых форм // Европейская рыжая полевка. М.: Наука, 1981. С. 87–97.
Ивантер Э. В., Моисеева Е. А. К экологии красной полевки (Clethrionomys rutilus Рall.) на юго-Западной периферии ареала // Труды Карельского научного центра РАН. 2015. № 1. С. 37–47.
Кошкина Т. В. Метод определения возраста рыжих полевок и опыт его применения // Зоологический журнал. 1955. Т. 34. Вып. 3. С. 631–639.
Коурова Т. П. Изменение рисунка жевательной поверхности зубов трех видов полевок на постювенильных стадиях развития // IV съезд Всесоюзного териологического общества: Тез. докл. М., 1986. Т. 2. С. 65.
Ларина Н. И., Еремина И. В. Каталог основных вариаций краниологических признаков у грызунов // Фенетика природных популяций. М.: Наука, 1988. С. 8–52.
Малеева А. Г. Об изменчивости зубов у полевок (Microtinae) // Эволюция грызунов и история формирования их современной фауны. Л.: Изд-во Зоол. ин-та АН СССР, 1976. С. 48–57.
Малеева А. Г., Шувалова Т. Г. Различные типы усложнения передней непарной петли М1, характерные для узкочерепной полевки // Фауна Урала и Европейского Севера. Свердловск: Уральский гос. ун-т, 1980. С. 5–14.
Маркова Е. А. Оценка сложности щечных зубов полевок (ARVICOLINAE, RODENTIA): ранжированный морфотипический подход // Зоологический журнал. 2013. Т. 92. № 8. С. 968–980.
Огнев С. И. Звери СССР и прилежащих стран. Грызуны . М.; Л.: Изд-во Академии наук СССР, 1940. Т. IV. 570 с.
Окулова Н. М., Андреева Т. А. Межвидовая и внутривидовая дифференциация лесных полевок рода Clethrionomys (Rodentia, Cricetidae) по данным изменчивости жевательной поверхности зуба М3 // Зоологический журнал. 2008. Т. 87. № 8. С. 991–1003.
Павлинов И. Я., Крускоп С. В., Варшавский А. А., Борисенко А. В. Наземные звери России: Справочник-определитель . М.: КМК, 2002. 304 с.
Поздняков А. А. Морфотипическая изменчивость жевательной поверхности коренных зубов серых полевок группы «maximowiczi» (Rodentia, Arvicolidae): опыт количественного статистического анализа // Зоологический журнал. 1993. Т. 72. Вып. 11. С. 114–125.
Смирнов Н. Г., Большаков В. Н., Бородин А. В. Плейстоценовые грызуны севера Западной Сибири . М.: Наука, 1986. 145 с.
Сорокина Н. В. Морфотипическая изменчивость структуры жевательной поверхности зуба М3 красной полевки (Clethrionomys rutilus Pallas, 1779) (RODENTIA, CRICETIDAE) северной и средней тайги Тюменской области // Известия Самарского научного центра Российской академии наук. 2011. Т. 13. № 1 (5). С. 1154–1157.
Сорокина Н. В., Сидорова С. К. Морфотипическая изменчивость структуры жевательной поверхности зубов М1 и М3 полевок р. Clethrionomys (Rodentia, Cricetidae) Среднего Зауралья // Известия Самарского научного центра Российской академии наук. 2016. Т. 18. № 2. С. 498–501.
Ямборко А. В. Популяционная экология лесных полевок (рода Clethtionomys) Северо-Восточной Азии : Автореф. дис. ... канд. биол. наук. Владивосток: Биолого-почвенный институт ДВО РАН, 2015. 24 с.
Chaline J., Laurin P., Brunet-Lecjmte P., Vitiot L. Morphological trends and rates of evolution in arvicolids (Arvicolidae, Rodentia): Towards a punctuated equilibria/diselibria model // Quaternary International. 1993. Vol. 19. P. 27–39.
Souto-Lima R. B., Millien V. The influence of environmental factors on the morphology of red-backed voles Myodes gapperi (Rodentia, Arvicolinae) in Québec and Western Labrador // Biological Journal of the Linnean Society. 2014. Vol. 112 (1). P. 204–218. DOI: 10.1111/bij.12263
Runck A., Matocq M. D., Cook J. A. Historic hybridization and persistence of a novel mito-nuclear combination in red-backed voles (genus Myodes) // Evolutionary Biology. 2009. Vol. 9. P. 114. DOI: 10.1186/1471-2148-9-114

 © 2011 - 2026
© 2011 - 2026