



Введение
Индекс флуктуирующей асимметрии (ФА) – это популярный показатель, который характеризуется незначительным отклонением от нуля разности величин правой и левой частей гомологичного билатерально симметричного признака. Для определения величины ФА и стабильности/нестабильности развития используются мерные или счетные билатерально симметричные признаки с широким диапазоном реакции на воздействие стресс-факторов (Palmer, Strobeck, 2003; Баранов и др., 2015). К таким признакам относятся наиболее генотипически или фенотипически изменчивые признаки. У представителей рода Липа мерные билатерально симметричные признаки листовых пластин обладают высокой изменчивостью, что связано с видовыми специфическими особенностями. Известно, что стабильность развития (СР) – это способность организма компенсировать и снижать отклонение от нормального онтогенетического развития по определенному канализирующему пути (Debat, David, 2001; Lens et al., 2002; Klingenberg, 2016).
Флуктуирующая асимметрия представляет интерес как в экологическом отношении, так и в аспекте эволюции и таксономии растительных организмов. Представители семейств Березовые и Липовые широко используются для определения уровня стабильности развития (Зорина, Коросов, 2015; Зыков и др., 2015), однако причины, вызывающие флуктуационные изменения в морфологических структурах билатеральных органов, остаются изученными слабо. Использование древесного вида как индикатора стабильности развития в баллах сталкивается с трудностями, связанными с генотипической чистотой используемого вида, популяционной гетерогенностью, влиянием факторов, включая освещение, водно-солевой состав и режим почвы, рельеф местности, другие физико-химические и иные особенности среды.
Методологически определение уровня стабильности развития вызывает вопросы. Методы измерения и подготовка гербарного материала занимают важную часть и подразумевают «слепой» непредвзятый подход (Kozlov, Niemelä, 2003; Коzlov, Zvereva, 2015). Традиционными методами, например нормализующей разности, предполагается работать с использованием отдельно взятых признаков, с нормальным распределением разности между правыми и левыми значениями. Такой подход по-прежнему находит применение, хотя оценка по одному-двум или множеству признаков зачастую не обеспечивает репрезентативной регистрации сигнала в виде повышенного значения ФА. Быстро развивающийся метод геометрической морфометрии позволяет дать более гармоничную интегративную оценку, приближая исследователей к истинным причинам повышения или снижения ФА. Развитие эпигенетики повлекло ряд новых исследований. Например, работы, проведенные над несколькими генерациями, указывают на генотипическую обусловленность процессов снижения – повышения величины ФА (Clarke, 1993; Freeman et al., 1993).
Остается признать, что «здоровье среды» – термин, с трудом прокладывающий дорогу в биоиндикации. Это мнение вытекает из результатов многочисленных работ в области динамики величины ФА, моделирующих экспериментов in vitro, сравнительных исследований по ФА разных видов, популяций, работ с отрицательными результатами (Zverev et al., 2018), наконец, работ с обоснованием парадоксальных эффектов влияния дозы стресса на величину ФА и стабильность развития (Erofeeva, 2015). Так, исследования ФА и СР в зависимости от градиента загрязнения техногенными токсинами показывают противоречивые результаты: как обратную (Kaligarič et al., 2008, Gostin, 2015), прямую связь (Беляева, 2013; Собчак и др., 2013; Koroteeva et al., 2015; Ivanov et al., 2015), так и неопределенные или условно-положительные результаты (Leung, 1996; Lung, 2000). В то же время изучение ФА листовых пластин в условиях интактных экосистем показало серьезное влияние высоты рельефа и других абиотических (Zvereva et al., 1997; Wuytack et al., 2011; Baranov, 2014а; Baranov, 2014b) и биотических факторов (Zvereva, 1997; Kozlov et al., 2017).
Влияние климатических факторов подробно изучено на примере дуба (Hódar, 2002), сосны обыкновенной (Kozlov, Niemelä, 2003), березы белой (Valkama, Kozlov, 2001) и некоторых травянистых растений (Alados et al., 2001). Совместное действие многих факторов, например влажности и пониженной температуры, остается невыясненным, поэтому эти факторы на фоне антропогенного стресса представляют интерес. Логическое продолжение исследований – фоновый мониторинг, изучение этапов морфогенеза листовых пластин (Anastasia, 2014) и работы в области эволюционного развития по определенному, канализирующему пути. Последний термин ассоциируют со свойством пластичности развития (developmental plasticity; Graham, 2010). В русскоязычной литературе продолжают использоваться привычные термины – фенотипическая или экологическая пластичность. Известно, что экологическая пластичность растений обуславливается буферной емкостью морфологических структур, что позволяет им активно адаптироваться к условиям среды. Вопрос о связи между пластичностью и стабильностью развития продолжает обсуждаться. Одновременно дискутируется вопрос о флуктуирующей асимметрии как об адаптационном сигнале, проявляющемся в траектории популяционного развития (Clarke, 1993; Debat, David, 2001; Graham et al., 2010). Модулярная концепция экологической пластичности подробно изложена во многих источниках, например в работе De Kroon (De Kroon et al., 2005). Согласно этой концепции норма реакции, как основная экологическая характеристика организма, охватывает сумму модулей пластичности корня, листа и стебля. Эволюция и адаптационные свойства пластической изменчивости (ПИ) в условиях пертурбации среды описаны также во многих обзорных работах (Lande, 2009; Gilbert et al., 2015; Abley et al., 2016), включая палеонтологические исследования размера и формы листовых пластин на примере подвидов клена в градиенте температурных условий (Royer et al., 2009). Устоявшимся мнением является корреляционная связь между стабильностью развития, фенотипической пластичностью и приспособленностью на генетическом уровне (Tonsor et al., 2013). Следовательно, пластическая изменчивость представляет интерес как спутник флуктуационной изменчивости как при антропогенном стрессе, так и при жестких климатических условиях. Несмотря на высокую вариацию признаков листьев липы и частое присутствие направленной асимметрии, мешающей определению ФА, по сравнению с видами р. Betula, представители рода Липа более удобны благодаря более стабильному хромосомному кариотипу. Известно, что близкие виды отличаются по величине ФА в ответ на содержание углекислого газа в среде и выбросов индустриальных предприятий, что подтверждает генотипическую основу стабильности развития (Hochwender, Fritz, 1999; Зорина, Коросов, 2015; Koroteeva et al., 2015).
Цель настоящей работы – определение стабильности развития и фенотипической пластической изменчивости у двух близких видов в двух урбанизированных стациях с разной степенью антропогенной нагрузки. В задачи входило исследование влияния климато-температурного фактора на стабильность развития и на пластическую изменчивость листовых пластин близких видов р. Липа. Проверяемая гипотеза заключалась в следующем: флуктуационная изменчивость связана с пластической изменчивостью, но у разных видов эта связь проявляется неодинаково, что объясняется генотипическими особенностями вида.
Материалы
Места сбора и подготовка гербарного материала
Известно, что липа широколистная (Tilia platyphyllos Scop.) – более эврибионтный вид по отношению к почвенным и климатическим условиям. Оба вида одинаково требовательны к освещению, что удобно для изучения генотипических и фенотипических свойств в контексте асимметрии и пластической изменчивости листовых пластин. Популяции обоих видов произрастают в восточной части Москвы, в Измайловском парке и на Измайловском бульваре. Природно-исторический парк «Измайлово» (55°47′N; 37°47′E) – это биоценоз с более чем 500 видами растений. Исходя из этого выбирались особи деревьев из различных частей парка, т. е. из различных фитоценозов.
Измайловский бульвар (55°48'13.8''N; 38°58'23.8''E) – это центр оживленной автострады, площадь стации на бульваре занимала около 5 кв км. Характерными свойстами данной стации были равномерность освещения, аэрации, отсутствие конкуренции и высокий уровень загрязнения атмосферы оксидами углерода, азота, серы и углеводородами.
Сбор листовых пластин проводился в генеративной стадии развития деревьев в 2016–2018 гг. В каждой стации листовые пластины шириной 3–4 см равномерно собирались с нижних частей крон десяти одновозрастных деревьев, по 10 образцов с каждого из 10 деревьев. Для измерения использовались признаки, изначально разработанные лабораторией В. М. Захарова (Захаров, Чубинишвили, 2001). Как показали предыдущие исследования (Баранов и др., 2015; Зыков и др., 2015), угловой признак был явно неудобен для измерения из-за высокой степени кривизны первой латеральной жилки. Он был заменен на альтернативный ему – линейный: расстояние между основанием второй жилки 1-го порядка и основанием первой жилки 2-го порядка на первой жилке 1-го порядка (рис. 1).
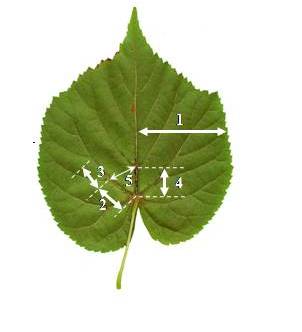
Рис. 1. Признаки листовой пластины, используемые для определения флуктуирующей асимметрии видов рода Tilia L.: 1 – ширина середины листа; 2 – расстояние между основаниями первой жилки 1-го порядка и второй жилки 2-го порядка; 3 – расстояние между основаниями второй и третьей жилок 2-го порядка; 4 – расстояние между основаниями первой и второй жилок 1-го порядка; 5 – расстояние между основанием второй жилки 1-го порядка и основанием первой жилки 2-го порядка на первой жилке 1-го порядка
Fig. 1. The traits used to determine the fluctuating asymmetry of species of genera Tilia L.: 1) the width of sheet mean; 2) distance between the bases of the first vein of the 1st order and the second vein of the 2nd order; 3) distance between the bases of the second and third veins of the 2nd order; 4) distance between the bases of the first and second veins of the 1st order; 5) distance between the base of the second vein of the 1st order and the base of the first vein of the 2nd order on the first vein of the 1st order
Листья высушивались под бумажным прессом, и через 2–3 дня проводились измерения мерной линейкой с точностью до 0.5 мм (ГОСТ 427-75), данные сохранялись в таблицах Excel. Трехкратное измерение показало крайне низкую ошибку, менее 1 % от величины ФА, что считается допустимым при тестировании асимметрии (Palmer, Strobeck, 2003). Принимая во внимание данные Velicković (Velicković, 2010) о наибольшей чувствительности признака длины листа и предыдущие результаты, в которых наиболее подходящим был другой признак (№ 4), мы решили использовать весь набор признаков, что представляется как преимущество интегративного подхода.
Методы
Подготовительный скрининг значений L и R (величины левого и правого признаков) играет существенную роль в тестировании ФА листовых пластин, не менее важную, чем подготовка гербарного материала и повышение точности измерения. Сначала была проведена инспекция данных на присутствие явно асимметричных признаков, например, если L и R различались более чем 8–10 раз, то такие (единичные) данные выбраковывались. Затем проводилась проверка на нормальность распределения (L – R) тестом Колмогорова – Смирнова с поправкой Лиллиефорса, т. к. эта разность и является отправной точкой тестирования ФА. Присутствие антисимметрии проверялось по эксцессу значений (L – R), т. к. величина эксцесса γ < –2 свидетельствует об отклонении от нормального распределения и о присутствии антисимметрии как возможном виде билатеральной асимметрии, который влияет на величину ФА. Направленная асимметрия (НА) определялась t-тестом с нулевой гипотезой H0: (L – R) = 0.
Принимая во внимание возможную связь между величиной признака (L + R)/2 и флуктуирующей асимметрией, проводился корреляционный анализ парных значений по Спирмену (Palmer, Strobeck, 2003). Тестирование статистической значимости факторов, влияющих на флуктуационную и пластическую изменчивость, проводилось факториальным дисперсионным анализом. Пластическая изменчивость определялась с помощью формулы PL = 1 – х/Х, где PL – величина пластической изменчивости, а х и Х соответствовали минимальному и максимальному значению величины признака (L + R)/2 листовой пластины с каждого дерева (Bruschi, 2003). Были использованы средние значения ПИ для каждого дерева, т. е. особь играла роль экспериментальной единицы (n = 10). Большая часть статистических анализов проводилась в программе STATISTICA10 (StatSoft Ink) с использованием уровня статистической значимости α = 0.05 %.
Результаты
Погодные условия
Вегетативный сезон 2017 г. был более холодный и влажный. Средняя температура в мае – августе 2017 г. была на 16–18 % ниже, а влажность на 2–3 % выше по сравнению с 2016 и 2018 гг. По данным Всероссийского НИИ гидрометеорологической информации, май 2017 г. был самым холодным с начала ΧΧΙ в., а июнь – самым прохладным за последние 70 лет (табл. 1).
Таблица 1. Климатические показатели, 2016–2018 гг.
Table 2. Climatic data, 2016–2018
| 2016 | 2017 | 2018 | 2016 | 2017 | 2018 | ||
| H | T | ||||||
| Май | 60.30 | 60.21 | 57.96 | 15.03 | 10.91 | 16.19 | |
| Июнь | 62.36 | 69.20 | 58.87 | 18.19 | 14.45 | 16.32 | |
| Июль | 69.54 | 73.51 | 72.63 | 20.96 | 17.94 | 20.47 | |
| Август | 72.92 | 69.82 | 63.51 | 19.48 | 18.84 | 19.64 | |
| Среднее | 66.28 | 68.18 | 63.24 | 18.41 | 15.53 | 18.15 | |
Примечание. T – средняя температура воздуха по Цельсию на высоте 2 м над уровнем земли; H – относительная влажность в % на высоте 2 м над уровнем земли.
Notes. T – average air temperature in Celsius at a height of 2 m above ground level; H – relative humidity in% at a height of 2 m above ground level.
Средняя температура воздуха в мае была ниже климатической нормы более чем на три градуса, были отмечены понижение температуры ниже нуля и появление снега в мае и июне, что влияет на фазы меристематической активности с последующей бутонизацией, облиствением и началом цветения липы в условиях России (Кищенко, 2015; Ерофеева, 2011; Уфимцева, Терехина, 2017).
Проверка первичных данных
Выборки (L – R) отклонялись от нормального распределения, поэтому для определения флуктуирующей асимметрии использовалась формула |Log L – Log R|. После нормализации только две выборки не удовлетворяли условиям нормального распределения и показали направленную асимметрию (t-test; р = 0.01). Учитывая невысокое отклонение от критериального t, было решено включить эти данные в исследование. В 90 % выборок (L – R) величина эксцесса γ была выше нуля, в остальных – эксцесс был меньше нуля, но не менее –0.2. Критическое значение γ, указывающее на антисимметрию, равно –0.68, при объеме наблюдений n = 100, следовательно, в выборках (L – R) антисимметрия выявлена не была. Парные коэффициенты корреляции, показывающие силу связи между величиной признака и ФА, показали у Tilia cordata слабую корреляционную зависимость (r = –0.11 ÷ 0.1; p < 0.05). Слабая корреляционная зависимость между величинами ФА признаков позволила сделать вывод о независимости признаков и пригодности их для определения стабильности развития. Влияние на величину ФА различных факторов показано в табл. 2.
Таблица 2. Влияние вида, года, места сбора и совместного действия факторов на величину ФА
Table 2. Effect of species, year, gathering place and combined effect of factors on FA value
| Источники вариации | SS | df | MS | F | p |
| Вид | 0.161 | 1 | 0.161 | 149.77 | 0.000 |
| Год сбора | 0.016 | 2 | 0.008 | 7.62 | 0.001 |
| Вид × год сбора | 0.012 | 2 | 0.006 | 5.79 | 0.003 |
| Место сбора | 0.003 | 1 | 0.003 | 2.74 | 0.098 |
| Вид × место сбора | 0.005 | 1 | 0.005 | 4.31 | 0.038 |
| Ошибка | 1.168 | 943 | 0.001 |
Примечание. Условные обозначения: SS – сумма квадратов; df – степень свободы; MS – средний квадрат; F – критерий Фишера; p – уровень статистической значимости.
Notes. SS – sum square; df – degree of freedom; MS – mean square; F – criterion of Fisher; p – level of statistical significance.
Фактически величина ФА зависела от вида, года сбора листовых пластин, но не зависела от расположения популяций.
Флуктуационная изменчивость
Как показано на рис. 2, на бульваре значение ФА только одного вида – липы плосколистной – было значительно выше по сравнению с контролем.
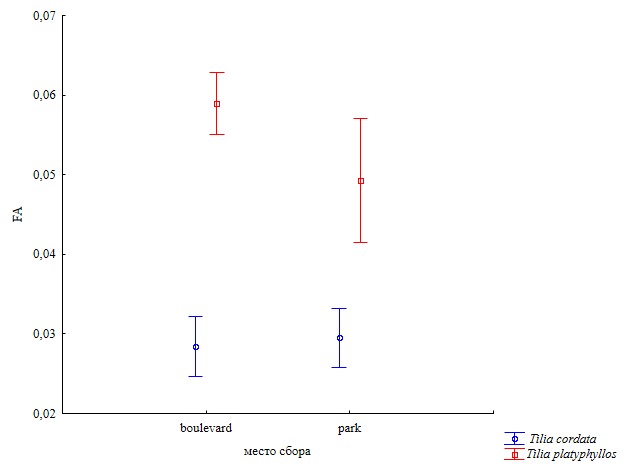
Рис. 2. Зависимость ФА от места сбора, F (1;945) = 4.31; p = 0.04. Вертикальные отрезки означают 95%-й доверительный интервал
Fig. 2. Dependence of FA on gathering place, F (1;945) = 4.31; p = 0.04. Vertical segments mean a 95 % confidence interval
Таким образом, липа плосколистная обладала флуктуационной изменчивостью, объясняемой действием 2 факторов: годом сбора и местом сбора материала. Наблюдения в 2017 г. выявили критические величины ФА только для липы плосколистной (рис. 3).
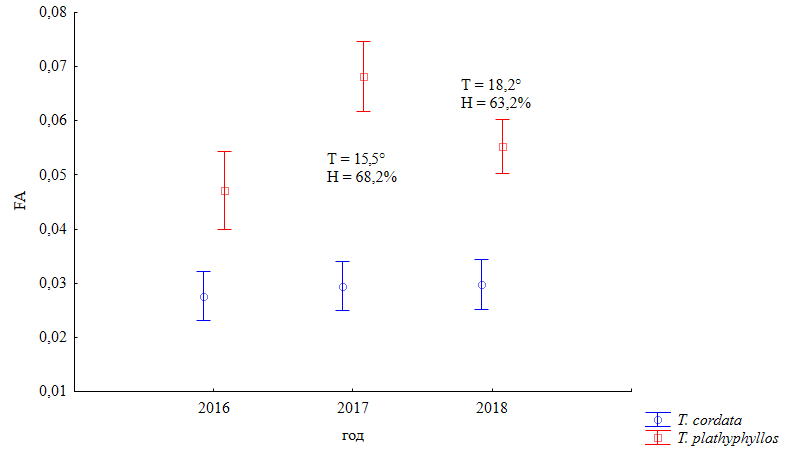
Рис. 3. Зависимость ФА от года сбора (в поле диаграммы – средняя годовая температура и влажность, среднее значение за май – август). F (2;943) = 5.79; p = 0.003. Вертикальные отрезки означают 95%-й доверительный интервал
Fig. 3. Dependence of FA on the year of collection (in the field of the diagram – average annual temperature and humidity, the average values for May – August). F (2;943) = 5.79; p = 0.003. Vertical segments mean a 95 % confidence interval
Пластическая изменчивость
Значимыми факторами, влиявшими на пластическую изменчивость листовых пластин, были видовая принадлежность и год сбора пластин (табл. 3).
Таблица 3. Влияние факторов на пластическую изменчивость
Table 3. Effect of some factors on phenotypic plasticity
| Источники вариации | SS | df | MS | F | p |
| Вид | 0.037 | 1 | 0.037 | 12.351 | 0.001 |
| Год сбора | 0.053 | 2 | 0.027 | 8.780 | 0.000 |
| Место сбора | 0.001 | 1 | 0.001 | 0.403 | 0.527 |
| Вид × год сбора | 0.039 | 2 | 0.019 | 6.412 | 0.003 |
| Вид × место сбора | 0.016 | 1 | 0.016 | 4.501 | 0.037 |
| Ошибка | 0.58 | 178 | 0.007 |
Примечание. Условные обозначения: SS – сумма квадратов; df – степень свободы; MS – средний квадрат; F – критерий Фишера; p – уровень статистической значимости.
Notes. SS – sum square; df – degree of freedom; MS – mean square; F – criterion of Fisher; p – level of statistical significance.
Пластическая изменчивость была неодинакова у двух видов (df = 62; p = 0.0002) и зависела от года сбора (df = 62; p = 0.006). У липы широколистной показатели ПИ, как и ФА, были выше, чем у липы мелколистной (p < 0.05; рис. 4).
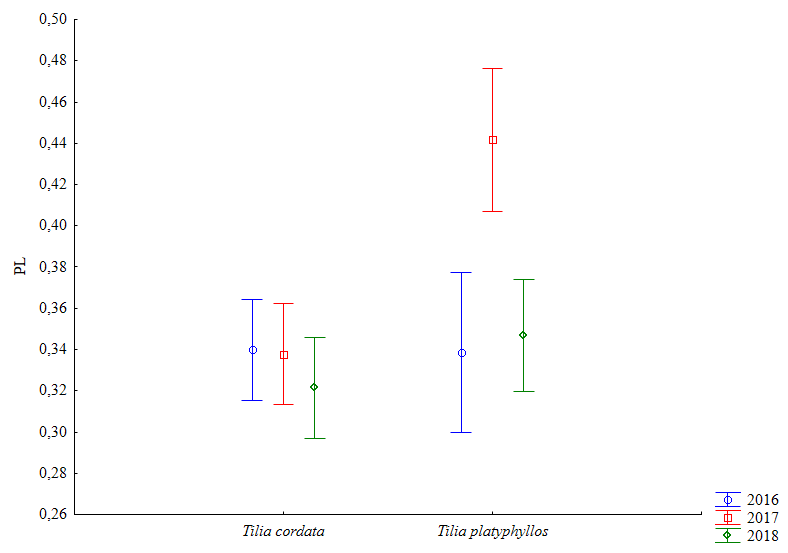
Рис. 4. Пластическая изменчивость, F (2;88) = 6.41; p = 0.003. Вертикальные отрезки означают 95%-й доверительный интервал.
Fig. 4. Plastic variation, F (2;88) = 6.41; p = 0.003. Vertical segments mean a 95 % confidence interval
2017 г. привел к повышению ФА только у липы широколистной, в этот год листовая пластина была значительно меньше: 44.34 ± 0.83 – в 2017 г.; 48.74 ± 1.04 – в 2016 г. и 46.85 ± 0.62 – в 2018 г. (р < 0.001). Не было получено статистически значимой корреляционной связи между ПИ и средней величиной признака. Характеристики листовой пластины липы мелколистной оставались инвариантными в течение всего срока наблюдения.
Верификация результатов
Поскольку листовые пластины были неодинаковы по величине (около 30 % выборок), то в задачу верификации входило определить, существенно ли изменялась стандартная ошибка ME при определении ФА в зависимости от величины признака. Если связь «ФА – признак» была сильнее, чем связь «ошибка – признак», то это давало право положительно верифицировать статистическую значимость величины ФА (Palmer, Strobeck, 2003). Был проведен регрессионный анализ с оценкой коэффициента корреляции Пирсона, т. к. выборки, содержащие средние значения на дерево, обладали нормальным распределением. Отношение разности корреляционных коэффициентов к ошибке измерения ФА было проверено в t-тесте исходя из предположения о нормальности распределения такого отношения:
ts = (rFA – rME)/ SEFA,
где ts – статистический критерий различия коэффициентов корреляции; rFA – значение r корреляционной связи между ФА и (R+ L)/2; rME – значение r корреляционной связи между ФА и ME; SEFA – стандартная ошибка ФА.
Для обоих видов статистическое значение критерия ts превышало табличное критериальное значение ts = 2.02; n = 40 (липа мелколистная – 62.4, липа широколистная – 43.4). Таким образом, влияние ошибки измерения на величину ФА на уровне индивидуальной изменчивости было несущественно. Обобщенная линейная модель (GLM) не показала различия в полученной зависимости в выборках, сгруппированных по годам сбора или по месту сбора (р > 0.05), т. е. в разные годы и в разных популяциях ошибка измерения одинаково не влияла на ФА. Описанную проверку статистической значимости стандартной ошибки авторы считают принципиальной в случае работы с образцами пластин разной величины, что встречается нередко в подобных исследованиях.
Обсуждение
Выбор отдельных признаков для определения стабильности развития в противоположность интегративному пути вызывает определенное сомнение и может быть оправдан, только если существует точная оценка градиента действия факторов на величину ФА признака (Baranov, 2014b; Баранов и др., 2015, Зыков и др., 2015). В нашем случае нормализация с применением логарифмирования позволяла оперировать нормально распределенными величинами у всех 5 признаков, что можно рассматривать, скорее, как удачу. В то же время сохраняется интрига использовать отдельные признаки (один-два), наиболее чувствительные к поллютантам (Baranov, 2014а; Koroteeva et al., 2015). Известно, что пластичность растений обуславливается буферной емкостью морфологических структур, которая позволяет им активно адаптироваться к условиям среды (Debat, David, 2001; Klingenberg, 2003; Tonsor et al., 2013; Venâncio et al., 2016). Следовательно, стрессовое климатическое воздействие и химическое загрязнение на бульваре влияли на ПИ, адаптируя растение. Известно, что на Кольском полуострове в холодное лето происходило увеличение ФА листьев березы белой, при этом температура являлась более значимым фактором, чем загрязнение диоксидом серы и никелем (Valkama, Kozlov, 2001). Как и в случае березы, климатический фактор играл более важную роль, чем действие химических поллютантов.
Липа широколистная – более теплолюбивый вид, с более выраженной стенобионтностью к климатическим факторам и физико-химическим свойствам почвы. Причину повышения ФА листовых пластин мы видим в снижении роста пластин и снижении активности фотосинтеза во время холодного лета. Сказанное подтверждается индикаторными значениями теплолюбивой липы плосколистной, которые смещены в правую положительную сторону (8–13) по сравнению с липой мелколистной (6–12 по шкале Цыганова).
Адаптацию липы широколистной к условиям среднерусской равнины авторы относят к одной из фаз реакции на стресс с повышением ФА листовых пластин. Следовательно, была подтверждена гипотеза о связи 2 видов изменчивости. Видовую генотипическую особенность этого интродуцента в виде повышенной пластической и флуктуационной изменчивости мы рассматриваем как «плату» за адаптацию в условиях климатических стрессовых факторов.
Заключение
Фенотипическая пластичность оказывает влияние на флуктуационную изменчивость (Houle, 2000; Sultan, 2003; Scheine, 2004; Lajus, Alekseev, 2003; Tucić et al., 2018). Стабильность развития представляется категорией, объясняющей процессы, обусловленные особенностями генотипа и внешней среды. ПИ – необязательный показатель, связанный корреляционной связью с ФА, но частое проявление, сопутствующее ФА. В данном случае повышенная пластическая изменчивость была фенотипическим отклонением, вызванным низкими температурами воздуха, а повышенная флуктуирующая асимметрия обуславливалась малым размером листовых пластин. Растущая пластина, как известно, развивается пульсирующе то в правом, то в левом направлении и обладает повышенной асимметричностью (Freeman et al., 1993). Известно, что атрибутом дестабилизации развития должно быть колебание дисперсии разности (L – R) в рамках одной величины признака (Palmer, Strobeck, 2003). В нашем случае пластичность развития листовых пластин была повышена, тем не менее ФА, хотя и повышенная, не являлась свидетельством снижения стабильности развития как отклонения в гомеостазе развития на биохимическом уровне (Klingenberg, 2003; Graham et al., 2010; Klingenberg, 2016).
Повышенная пластическая изменчивость в листовых пластинах липы плосколистной как видоспецифическое свойство, без дисбаланса в гомеостазе развития, должно учитываться в исследованиях, связанных с долгосрочным мониторингом стабильности развития древесных растений.
Библиография
Баранов С. Г., Зыков И. Е., Федорова Л. В. Изучение внутривидовой изменчивости липы мелколистной (Tilia cordata Mill.) на основе билатеральной асимметрии листовых пластин // Вестник Томского государственного университета. Биология. 2015. № 2 (30) C. 134–145.
Беляева Ю. В. Показатели флуктуирующей асимметрии Betula pendula Roth. в условиях антропогенного воздействия (на примере г. Тольятти) // Известия Самарского научного центра Российской академии наук. 2013. № 15. С. 3–7.
Ерофеева Е. А. Влияние автотранспортного загрязнения на скорость выхода из состояния зимнего покоя и окончание вегетации у липы мелколистной // Вестник ННГУ. 2011. № 2-2. URL: https://cyberleninka.ru/article/n/vliyanie-avtotransportnogo-zagryazneniya-na-skorost-vyhoda-iz-sostoyaniya-zimnego-pokoya-i-okonchanie-vegetatsii-u-lipy-melkolistnoy (дата обращения: 25.08.2018).
Захаров В. М., Чубинишвили А. Т. Мониторинг здоровья среды на охраняемых природных территориях . М.: Центр экологической политики России, 2001. 148 с.
Зорина А. А., Коросов А. В. Характеристика флуктуирующей асимметрии листа двух видов берез в Карелии // Труды Карельского научного центра Российской академии наук. 2007. Вып. 11. С. 28–36
Зыков И. Е., Федорова Л. В., Баранов С. Г. Оценка биоиндикационного значения уровня изменчивости параметров листовых пластинок липы мелколистной (Tilia cordata Mill.) в условиях города Орехово-Зуево и Орехово-Зуевского района // Вестник Московского государственного областного университета. Серия: Естественные науки. 2015. № 1. С. 15–21.
Кищенко И. Т. Рост и развитие интродуцированных видов рода Tilia L. (tiliaceae) в условиях Карелии // Arctic Evironmental Research. 2015. № 4. URL: https://cyberleninka.ru/article/n/rost-i-razvitie-introdutsirovannyh-vidov-roda-tilia-l-tiliaceae-v-usloviyah-karelii (дата обращения: 25.08.2018).
Собчак Р. О., Афанасьева Т. Г., Копылов М. А. Оценка экологического состояния рекреационных зон методом флуктуирующей асимметрии листьев Betula pendula Roth. // Вестник Томского государственного университета. 2013. № 368. С. 195–199.
Уфимцева М. Д., Терехина Н. В. Оценка экологического состояния Центрального района Санкт-Петербурга на основе экофитоиндикации // Вестник СПбГУ. Науки о Земле. 2017. № 2. URL: https://cyberleninka.ru/article/n/otsenka-ekologicheskogo-sostoyaniya-tsentralnogo-rayona-sankt-peterburga-na-osnove-ekofitoindikatsii (дата обращения: 25.08.2018).
Abley K., Locke J. C., Leyser H. O. Developmental mechanisms underlying variable, invariant and plastic phenotypes // Annals of botany. 2016. Apr. 1. Vol. 117 (5). P. 733–748.
Alados C. L., Navarro T., Escós J., Cabezudo B., Emlen J. M. Translational and fluctuating asymmetry as tools to detect stress in stress-adapted and nonadapted plants // International Journal of Plant Sciences. 2001. May. Vol. 162 (3). P. 607–616.
Anastasia Z. The formation of fluctuating asymmetry during individual development of Вetula pendula // Principy Èkologii. 2014. Vol. 4. Jan. 1. P. 31–52.
Baranov S. G. Littleleaf Linden Tilia cordata (Mill.): Only Some Bilateral Traits Indicate Chemical Pollution Induced by Chemical Plant // Advances in Biological Research. 2014а. Vol. 8 (4). P. 143–148.
Baranov S. G. Use of morphogeometric method for study fluctuating asymmetry in leaves Tilia cordata under industrial pollution // Adv. Environ. Biol. 2014b. Vol. 8 (7). P. 2391–2398.
Baranov S. G. Use of a geometric morphometric method to determine the developmental stability of Betula pendula Roth. // Biology Bulletin. 2017. Sep. 1. Vol. 44 (5). P. 547–551.
Baranov S. G. Use of MorphoJ Soft Package for Phenotypic and Genotypic Variety Testing (English Oak Case Study) // Russian Journal of Genetics: Applied Research. 2018. Vol. 8. No 1. P. 22–30.
Bruschi, P., Grossoni P., Bussotti F. Within-and among-tree variation in leaf morphology of Quercus petraea (Matt.) // Liebl. natural populations. Trees. 2003. Vol. 17 (2). P. 164–172.
Clarke G. M. Fluctuating asymmetry of invertebrate populations as a biological indicator of environmental quality // Environmental Pollution. 1993. Jan. 1. Vol. 82 (2). P. 207–211.
Cornelissen T., Stiling P., Drake B. Elevated CO2 decreases leaf fluctuating asymmetry and herbivory by leaf miners on two oak species // Global Change Biology. 2004. Jan. 1. Vol. 10 (1). P. 27–36. DOI: 10.1046/j.1529-8817.2003.00712.x.
De Kroon H., Huber H., Stuefer J. F., and Van Groenendael J. M. A modular concept of phenotypic plasticity in plants // New Phytol. 2005. Vol. 166. P. 73–82. DOI: 10.1111/j.1469-8137.2004.01310.x.
Debat V., David P. Mapping phenotypes: canalization, plasticity and developmental stability // Trends in Ecology & Evolution. 2001. Oct. 1. Vol. 16 (10). P. 555–561. DOI: org/10.1016/S0169-5347(01)02266-2.
Erofeeva E. A. Hormesis and paradoxical effects of Drooping Birch (Betula pendula Roth) parameters under motor traffic pollution // Dose-Response. 2015. Jun. 1. Vol. 13 (2). DOI: 10.1177/1559325815588508.
Freeman D. С., Graham J. H., Emlen J. M. Developmental stability in plants: symmetries, stress and epigenesist // Genetica. 1993. No 89. P. 97–119.
Gilbert S. F., Bosch T. C., Ledón-Rettig C. Eco-Evo-Devo: developmental symbiosis and developmental plasticity as evolutionary agents // Nature Reviews Genetics. 2015. Oct. Vol. 16 (10). P. 611.
Gostin I. Air pollution stress and plant response // Plant Responses to Air Pollution. 2016. P. 99–117.
Graham J. H., Raz S., Hel-Or H., Nevo E. Fluctuating Asymmetry: Methods, Theory, and Applications // Symmetry. 2010. Vol. 2. P. 466–540. DOI: 10.3390/sym2020466.
Hochwender C. G., Fritz R. S. Fluctuating asymmetry in a Salix hybrid system: the importance of genetic versus environmental causes // Evolution. 1999. Apr. Vol. 53 (2). P. 408–416.
Hódar J. A. Leaf fluctuating asymmetry of Holm oak in response to drought under contrasting climatic conditions // Journal of arid environments. 2002. Oct. 1. Vol. 52 (2) P. 233–243.
Houle D. A simple model of the relationship between asymmetry and developmental stability // Journal of Evolutionary Biology. 13.4.2000. P. 720–730. Instab.: Causes and Consequences. Polak M, editor. Oxford University Press, 2003. DOI: 10.1046/j.1420-9101.2000.00195.x.
Ivanov V. P., Ivanov Y. V., Marchenko S. I., Kuznetsov V. V. Application of fluctuating asymmetry indexes of silver birch leaves for diagnostics of plant communities under technogenic pollution // Russian journal of plant physiology. 2015. May 1. Vol. 62 (3). P. 340–348.
Kaligarič M., Tognetti R., Janžekovič F., Raschi A. Leaf Fluctuating Asymmetry of Myrtus Communis L., Affected by Increases in Atmospheric CO2 Concentration: Evidence from a Natural CO2 Spring // Polish Journal of Environmental Studies. 2008. Jul. 1. Vol. 17 (4).
Klingenberg C. P. A developmental perspective on developmental instability: theory, models and mechanisms // Developmental instability: causes and consequences. 2003. P. 14–34.
Klingenberg, C. P., Duttke S., Whelan S., Kim M. Developmental plasticity, morphological variation and evolvability: A multilevel analysis of morphometric integration in the shape of compound leaves // J. Evol. Biol. 2012. Vol. 25. P. 115–129.
Klingenberg C. P. Size, shape, and form: concepts of allometry in geometric morphometrics // Development genes and evolution. 2016. Jun. 1. Vol. 226 (3). P. 113–137. DOI: 10.1007/s00427-016-0539-2.
Koroteeva E. V., Veselkin D. V., Kuyantseva N. B., Chashchina O. E. The size, but not the fluctuating asymmetry of the leaf, of silver birch changes under the gradient influence of emissions of the Karabash Copper Smelter Plant // Doklady Biological Sciences. 2015. Vol. 460. No 3. P. 364–367. DOI: 10.1134/S0012496615010056.
Kozlov M. V., Cornelissen T., Gavrikov D. E., Kunavin M. A., Lama A. D., Milligan J. R., Zverev V., Zvereva E. L. Reproducibility of fluctuating asymmetry measurements in plants: Sources of variation and implications for study design // Ecological indicators. 2017. Feb. 1. Vol. 73. P. 733–740/ DOI: 10.1016/j.ecolind.2016.10.033
Kozlov M. V., Niemelä P. Drought is more stressful for Northern populations of Scots pine than low summer temperatures // Silva Fennica. 2003. Vol. 37. P. 175–180.
Kozlov M. V., Zverev V., Zvereva E. L. Do defoliating insects distinguish between symmetric and asymmetric leaves within a plant? // Ecological Entomology. 2018. Vol. 43 (5). P. 656–664.
Kozlov M. V., Zvereva E. L. Confirmation bias in studies of fluctuating asymmetry // Ecological indicators. 2015. Oct. 1. Vol. 57. P. 293–297. DOI: 10.1016/j.ecolind.2015.05.014.
Lajus D. L., Alekseev V. R. Phenotypic variation and developmental instability of life-history traits: a theory and a case study on within-population variation of resting eggs formation in Daphnia // Journal of Limnology. 2004. Sep. 1. Vol. 63 (1). P. 37–44. DOI: 10.4081/jlimnol.2004.s1.37.
Lande R. Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation // J. Evol. Biol. 2009. Vol. 22. P. 1435–1446. DOI: 10.1111/j.1420-9101.2009.01754.x.
Lens L. U., Van Dongen S., Kark S. et al. Fluctuating asymmetry as an indicator of fitness: can we bridge the gap between studies? // Biological Reviews. 2002. Feb. Vol.77 (1). P. 27–38. DOI: 10.1017S1464793101005796.
Leung B., Forbes M. R. Fluctuating asymmetry in relation to stress and fitness: effects of trait type as revealed by meta-analysis // Ecoscience. 1996. Jan 1. Vol. 3 (4). P. 400–413. DOI: 10.1080/11956860.1996.11682357.
Leung B., Forbes M. R., Houle D. Fluctuating asymmetry as a bioindicator of stress: comparing efficacy of analyses involving multiple traits // The American Naturalist. 2000. Jan. Vol. 155 (1). P. 101–115.
Palmer A. R., Strobeck C. H. Fluctuating Asymmetry Analyses Revisited // Developmental Instab.: Causes and Consequences. Polak M, editor. Oxford University Press, 2003. P. 279–319.
Royer D. L., Meyerson L. A., Robertson K. M., and Adams J. M. Phenotypic plasticity of leaf shape along a temperature gradient in Acer rubrum // PLoS ONE. 2009. Vol. 4(10). DOI: 10.1371/journal.pone.0007653.
Scheiner S. M. The genetics of phenotypic plasticity. XIII. Interactions with developmental instability // Ecology and evolution. 2014. Apr. 1. Vol. 4 (8). P. 1347–1360. DOI: 10.1002/ece3.1039.
Sultan S. E. Phenotypic plasticity in plants: a case study in ecological development // Evolution & development. 2003. Jan. 1. Vol. 5 (1). P. 25–33. DOI: 10.1046/j.1525-142X.2003.03005.x.
Tonsor S. J., Elnaccash T. W., Scheiner S. M. Developmental instability is genetically correlated with phenotypic plasticity, constraining heritability, and fitness // Evolution. 2013. Oct. 1. Vol. 67 (10). P. 2923–2935. DOI: 10.1111/evo.12175.
Tucić B., Budečević S., Manitašević Jovanović S. et al. Phenotypic plasticity in response to environmental heterogeneity contributes to fluctuating asymmetry in plants: first empirical evidence // Journal of Evolutionary Biology. 2018. Vol. 31 (2). P. 197–210. DOI: 10.5061/dryad.8th5m.
Valkama J., Kozlov M. V. Impact of climatic factors on the developmental stability of mountain birch growing in a contaminated area // Journal of Applied Ecology. 2001. Vol. 38. P. 665–673.
Veličković M. V. Reduced developmental stability in Tilia cordata leaves: effects of disturbed environment // Periodicum biologorum. 2010. Sep. 1. Vol. 112 (3). P. 273–281. P. 296–303.
Venâncio Henrique, Estevao Alves-Silva and Jean Carlos Santos. Leaf phenotypic variation and developmental instability in relation to different light regimes // Acta Botanica Brasilica. 2016. Vol.30 (2). P. 296–303. DOI: 10.1590/0102-33062016abb0081
Wuytack T., Wuyts K., Van Dongen S., Baeten L., Kardel F., Verheyen K., Samson R. The effect of air pollution and other environmental stressors on leaf fluctuating asymmetry and specific leaf area of Salix alba L. // Environmental pollution. 2011. Oct. 1. Vol. 159 (10). P. 2405–2411.doi.org/10.1016/j.envpol.2011.06.037
Zverev V., Lama A. D., Kozlov M. V. Fluctuating asymmetry of birch leaves did not increase with pollution and drought stress in a controlled experiment // Ecological indicators. 2018. Jan. 31. Vol. 84. P. 283–289.DOI: 10.1016/j.ecolind.2017.08.058
Zvereva E. L., Kozlov M. V., Haukioja E. Stress responses of Salix borealis to pollution and defoliation // Journal of Applied Ecology. 1997. Vol. 34 (6) Dec. 1. P. 1387–1396. DOI: 10.2307/2405256.
Zvereva E. L., Kozlov M. V., Niemelä P., Haukioja E. Delayed induced resistance and increase in leaf fluctuating asymmetry as responses of Salix borealis to insect herbivory // Oecologia. 1997. Feb. 1. Vol. 109 (3). P. 368–373.
Благодарности
Авторы приносят благодарность сотрудникам Первого Московского государственного медицинского университета им. И. М. Сеченова за сбор и подготовку гербарного материала.

 © 2011 - 2024
© 2011 - 2024