



Введение
Выявление пространственно-типологической дифференциации различных биотических групп – одна из актуальных задач современной экологической биогеографии. Практическое использование полученной при этом информации лежит в сфере решения природоохранных проблем, а научно-познавательное значение состоит в детализации и конкретизации биогеографических представлений, в частности степени соответствия зонально-подзональных границ, интра- и экстразональных включений и территориальной неоднородности различных биотических групп, рассматриваемых по отдельности, а также экосистем в целом, сравниваемых на примере эталонных групп. Этим вопросам посвящена серия публикаций по территории Западно-Сибирской равнины, Алтайской и Кузнецко-Салаирской горных областей, упомянутых в библиографии к данной статье.
Объектом исследования считали статистические ансамбли с внешним ограничением (Сержантов, 1972). К ним отнесены группы (группировки) растений или животных, существующих на одной и той же территории и испытывающих одинаковый набор структурообразующих факторов среды и их неразделимых сочетаний – природно-антропогенных режимов. Из-за различий в толерантности к условиям среды указанных групп список факторов и режимов может существенно отличаться. Такой набор факторов и сочетаний выявлен для каждой группы отдельно с помощью кластерного и факторного анализа, а не задан априорно по сведениям, известным из предыдущих публикаций. Результатом процесса формирования территориальной неоднородности групп можно считать список таких факторов, минимизированный, но достаточно полный по информативности, измеренной через учтенную классификацией групп дисперсию матрицы их сходства.
Такие группы (группировки) мы называем сообществами, в соответствии с их определением, где сообщество – «это совокупность совместно обитающих организмов разных видов, представляющая собой определенное экологическое единство (например, фитопланктон какого-либо озера, почвенные животные участка леса). Иногда, сообщество… трактуется как биоценоз. Выделяют также сообщество растений (фитоценоз) и… животных (зооценоз)... Сообщество является элементом экосистемы (или биогеоценоза)» (Биологический энциклопедический словарь, 1986). Аналогичные определения приводит Н. Ф. Реймерс (1980, 1990). При этом термины «сообщество», «биота», «население» и «ансамбль» мы считаем синонимами.
Современную изученность указанных связей для лишайников, с учетом статистической оценки, нельзя считать достаточной. Так, в монографии Н. В. Седельниковой (2017) такие оценки отсутствуют, хотя по животному населению и растительности, как отдельно, так и в целом по группам, по исследованной территории они имеются (Равкин и др., 2007, 2008, 2009; Пространственно-типологическая дифференциация…, 2011).
Поэтому цель исследований, результаты которых изложены в предлагаемой вниманию читателя статье, сводится к описанию пространственно-типологической организации сообществ, то есть зависимости их изменчивости от среды. Задачи включают выявление изменчивости сообществ на указанной территории и факторов среды, коррелирующих с ней, а также оценку силы и общности их связи, как отдельно по каждому фактору или режиму, так и расчета множественной информативности всей их совокупности. Статьи такой направленности следует считать эколого-биогеографическими и относить к глобальной экологии. Мы далеки от мысли пересматривать зональное деление Палеарктики. Наши исследования преследуют другую цель: сопоставить неоднородность сообществ различных «групп» растений и животных при использовании одинаковых подходов, формализованных методов и на одной и той же территории, а также оценить степень отличия выявленных нами границ с зональными, которые проведены на основе умозрительных представлений по облику растительности.
Материалы
Исследования проведены на основе данных по лишайникам и лихенофильным грибам по монографии Н. В. Седельниковой (2017). Они включают сведения о распределении 1903 видов лишайников и лихенофильных грибов, часть из которых представлена внутривидовыми таксонами. В общей сложности в расчеты включены 1942 формы, встреченные в 61 биотопе равнинной и горной частей Западной Сибири (Западная Сибирь, 1963). Поймы, агро- и селитебные ландшафты не обследованы, поэтому карта и выполненные классификации лихенобиоты составлены по аналогии с представлениями о «восстановленной» растительности, а пойменные сообщества «генерализованы» и входят в комплексы окружающих ландшафтов. Биотопы выделены по эдификаторным видам растений и соответствуют выделам карты «Растительность Западно-Сибирской равнины» (1976) с учетом рельефа, заболоченности и поемности.
Автором монографии на основании многолетних исследований экспертно выделено семь групп ценотической значимости видов (по встречаемости и степени преобладания): 1 – доминирующие; 2 – содоминирующие; 3 – постоянные; 4 – сопутствующие; 5 – редкие; 6 – отсутствующие; 7 – информация по которым отсутствует. Для выполнения расчетов этим группам присвоены условные баллы значимости в порядке, обратном их перечислению, – от 7 до 3, а последним двум группам – 0, поскольку в них входят виды, не обнаруженные при проведении работ в указанных биотопах вследствие очень низкой встречаемости. В 5 из 66 обследованных биотопов лишайники не обнаружены. Поскольку нет стопроцентной гарантии, что в таких выделах лишайников в принципе нет, эти биотопы объединены с близкими по растительности, в которых встречен хотя бы один вид лишайников или лихенофильных грибов. Так, галофитные луга включены в группу галофитных сообществ, лесостепные верховые островные болота объединены с низинными той же подзоны, лесостепные луговые степи и остепненные луга – с луговыми степями в пределах степной зоны, а сухие настоящие – с остальными настоящими степями. Такое объединение призвано исключить из расчетов биотопы, где лишайники не встречены, поскольку наличие нулевых оценок встречаемости порождает при классификации полную неопределенность. Это обусловлено тем, что при нулевой оценке сходства варианты населения можно считать как полностью похожими, так и абсолютно непохожими. Дело в том, что бесконечно малая и бесконечно большая величина оценки, сопоставленная с нулевой, дает полную непохожесть коэффициентов сходства, хотя первая из них почти не отличается от нуля, а вторая – существенно иная. Именно это порождает в процессе классификации необъяснимые ситуации. Объединяя «нулевые» варианты с экологически близкими по условиям среды, можно избавиться от указанных противоречий, почти не теряя информации об отсутствии лишайников в подобных местообитаниях. После такого объединения расчеты проведены не по 66 исходным биотопам, а по 61.
Методы
Для сжатия (обобщения) информации о распределении лишайников и территориальной неоднородности их биоты, а также для выявления основных структурообразующих факторов среды в работе использованы нелинейные методы статистики и кластерный анализ. При описании структуры пространственно-типологической неоднородности биоты использован метод построения и анализа графов такой изменчивости. Структурные графы иллюстрируют смену вариантов биот по биотопам и позволяют выявлять основные тренды (длящиеся, а не только локальные изменения) по значимым связям между таксонами выполненных классификаций. Это способствует формированию более представительного набора структурообразующих факторов среды и природных режимов как неразделимых сочетаний таких факторов. Для оценки силы связи между неоднородностью лихенобиоты и коррелирующими с ней факторами среды применена линейная качественная аппроксимация матриц связи (один из методов регрессионного анализа; Равкин и др., 1978). Множественная оценка связи всех факторов и режимов с территориальной неоднородностью биоты позволяет судить о степени информативности излагаемых представлений и о скоррелированности факторов между собой. Графы представляют собой модификацию иерархической классификации с учетом не только внутригруппового сходства биотических сообществ, но и межгрупповой близости их. Выполненное для проверки шкалирование, в принципе, подтвердило правильность ориентации построенных графов в факторном пространстве. Факторы среды выявлены по результатам кластерного анализа. Например, если совокупность вариантов биоты разделена на три кластера, при этом в первый из них вошли лесные сообщества, во второй – территории, где облесенные участки чередуются с открытыми, а в третий – только открытые местообитания, можно утверждать, что неоднородность выборки совпадает с фактором облесенности. Ее можно представить в виде трех выделенных номинальных градаций: высокая, средняя и низкая облесенность. В дальнейшем эти экспертные градации могут быть использованы при оценке связи с этим фактором изменчивости сообществ.
Природные режимы также выявляют по результатам кластерного анализа, когда их образуют неразделимые сочетания факторов, например сообществ лесов северной тайги Западной Сибири или лесов средней тайги и южных подзон той же территории. Перечни таких режимов приведены на рисунках и в легенде карты. Все факторы и режимы оценены в номинальной шкале. Режимы, так же как и отдельно взятые факторы среды, используют для оценки силы и общности связи с помощью линейной качественной аппроксимации. Для оценки сходства видов лишайников по распространению использован коэффициент Жаккара – Наумова (для количественных признаков) с предварительной нормировкой показателей. При классификации по этим коэффициентам учитывают только наибольшие значения в пересечении по распределению. Для расчета сходства биот применен коэффициент корреляции, в котором исключена процедура центрирования. Он, так же как коэффициент Жаккара и Жаккара – Наумова, не имеет отрицательных значений, но в отличие от них учитывает сходство по отсутствию видов в неблагоприятных биотопах, что в данном случае дает дополнительную информацию о неоднородности лихенобиоты.
При недостаточной агрегации сообществ и видов после первой процедуры классификации на исходных данных проводили повторную агрегацию на матрицах межгрупповой связи (средних значениях коэффициентов сходства сообществ, вошедших в значимо сходные группы). Результаты первого разбиения всей совокупности сообществ считали делением на типы биоты, группы, выделенные в процессе доразбивки типов, – подтипами, а после повторной агрегации – надтипами. В дальнейшем, в результате анализа составленной обзорной карты выделены типолого-хорологические полосы – территории с одинаковым «рисунком», со сходным цветом и сочетанием выделов, преобладающих по площади и «чуждых» включений, занимающих меньшую площадь. По видам лишайников и лихенофильных грибов, из-за существенно меньшей общности, результаты первого разбиения считали делением на подтипы распределения, повторной агрегации – типами. В надтипы они объединены по сходству растительности на занятой ими территории.
Большая часть методов и методологических подходов описаны ранее (Куперштох и др., 1976; Куперштох, Трофимов, 1976; Равкин и др., 1978; Миркин, 1980; Равкин, Ливанов, 2008). В этих публикациях указаны разработчики использованного программного обеспечения и приведены ссылки на соответствующие работы.
Результаты
Классификация видов лишайников и лихенофильных грибов по сходству распределения
Автоматическая классификация видов по исходным данным показала возможность их группировки в 38 кластеров. При повторной агрегации они объединены в 14 групп, принятых в качестве типов распространения (рис. 1). Результаты территориального анализа этих групп по сходству биотопов, общих для отнесенных к одному типу видов, дали основание для выделения трех надтипов распространения лишайников: Тундрового, Степного и Лесного. Такое классификационное представление учитывает 36 % дисперсии матрицы сходства по 1942 формам лишайников. Еще информативнее деление на типы и надтипы с учетом межгруппового сходства – 49 % (множественный коэффициент корреляции – 0.70). В названиях надтипов распространения отражены характеристики преобладающих по значимости биотопов, где вошедшие в типовые группы виды совместно встречаются наиболее часто, судя по вкладу в коэффициент внутригруппового сходства. Следует отметить, что Степной и Лесной надтипы распространения через краевые группы значимо сходны между собой, в то время как Тундровый имеет с ними лишь запороговое сходство. Кроме того, в этот надтип распространения входят виды, слабо (хотя и максимально) сходные по их встречаемости в равнинных моховых тундрах, лесотундровых травяных ивняках и ольховниках, а также в арктических болотах (типы 1.4–1.6). В Лесной надтип включены виды, пересекающиеся в распространении в лесостепных сосняках и их производных – березово-сосновых и мелколиственных лесах (тип 3.4). Эти виды имеют лишь слабое запороговое сходство с группой видов горных лесов. Все степные типы распространения значимо связаны между собой, хотя и включают лишайники тундростепей.
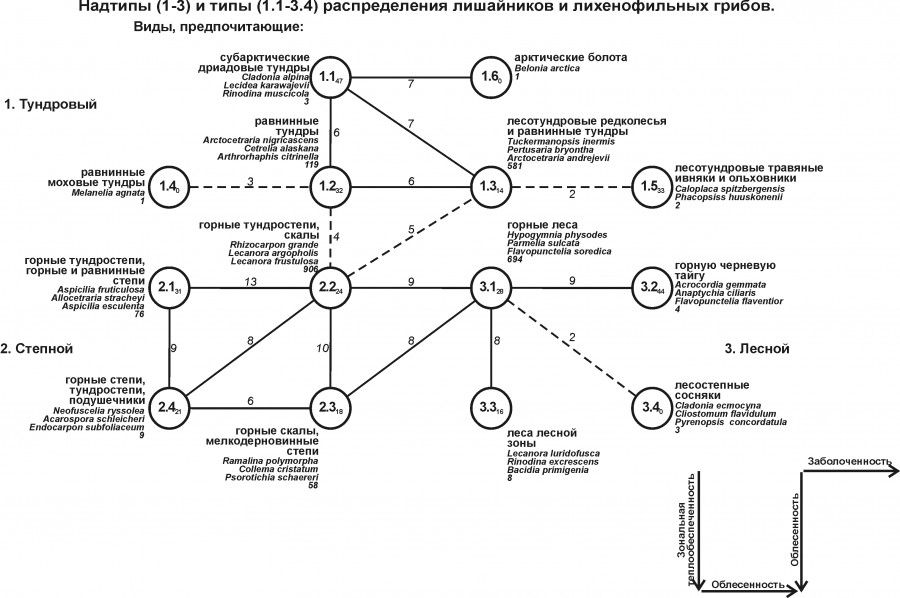
Рис. 1. Структура неоднородности в распределении лишайников и лихенофильных грибов Западной Сибири. Состав биотопов и первые три вида с наибольшей суммарной ценотической значимостью приведены рядом с номерами таксонов классификации. После перечисления лидеров указано общее число видов, отнесенных к данному типу распределения. При равной сумме показателей ценотической значимости по каждому виду в отдельности они перечислены в алфавитном порядке, сначала по названиям родов, а затем – видов. На рис. 1–3 сплошной чертой показаны сверхпороговые связи, прерывистой – максимальные при отсутствии значимых, пунктиром – вспомогательные. Рядом со связями приведены значения межгруппового сходства. Порог значимости соответствует наименьшей для каждого графа величине сверхпороговой связи. Индексом у номера таксона показано среднее внутригрупповое сходство. Стрелками обозначено направление увеличения влияния факторов среды, выявленных по графу
Fig. 1. Structure of heterogeneity in the lichen and lichenophilic fungi distribution of Western Siberia. The composition of biotopes and the first three species with the greatest total cenotic significance are listed next to the classification taxa numbers. After the leaders, the total number of species assigned to a given type of distribution is indicated. With an equal amount of cenotic significance indicators for each species individually, they are listed in alphabetical order, first by the names of the genera, and then by the species. In Fig. 1–3 a solid line shows superthreshold connections, intermittent – maximum ones in the absence of significant ones, dashed – auxiliary ones. Next to the links the values of intergroup similarity are presented. The threshold of significance corresponds to the smallest value of the threshold connection for each graph. The index of the taxon number shows the average intragroup similarity. Arrows indicate the direction of increasing the influence of environmental factors identified by the graph.е
Наиболее представительным по числу объединяемых видов следует считать Степной надтип распространения (всего 1049 форм; по отдельным типам – от 9 до 694). Менее представителен по этому показателю Лесной надтип (709; 3–694), и последнее место занимает Тундровый надтип (192; 1–119). Среди типов распространения наиболее представительна группа 2.2. К ней отнесены 906 форм, пересекающихся между собой по распределению в горных тундростепях и на скалах. На втором месте стоит тип лишайников горных лесов (694 вида), на третьем – виды равнинных тундр и лесотундровых редколесий (581). На четвертом месте – лишайники равнинных тундр (119). По остальным надтипам представленность в типовых группировках колеблется в пределах от одной до 76 форм.
Таким образом, можно утверждать, что в список основных факторов среды, коррелирующих с распространением лишайников на территориях со значимым пересечением по их встречаемости, входят тип растительности, зональная теплообеспеченность, рельеф и частично заболоченность.
Структура пространственно-типологической изменчивости лихенобиот
Первичная классификация 61 варианта лихенобиот показала целесообразность выделения 17 типов сообществ, объединенных в результате последующей агрегации в 8 надтипов (рис. 2). В первый надтип включены комплексы нивально-субнивальных и альпийско-субальпийских лугов, во второй – тундр, кустарников, редколесий, лесов (исключая степные сосняки) и болот (кроме лесостепных и степных). В третий надтип вошли сообщества лесных горных лугов, а в последующие – с четвертого по шестой – различные варианты степной биоты. При этом не исключено, что лихенобиота вторичных мелкодерновинных степей, образующих отдельный (пятый) надтип, вероятно, относится к четвертому надтипу, а выделение ее в отдельный таксон такого высокого ранга – случайно. По суммарной фитоценотической значимости и числу отмеченных видов наиболее богат второй (Тундрово-лесной) надтип биоты, на втором месте – шестой биотический надтип. Менее богаты надтипы 4, 1 и 3, т. е. сообщества лугов и луговых степей (последние – с включением степных сосняков).
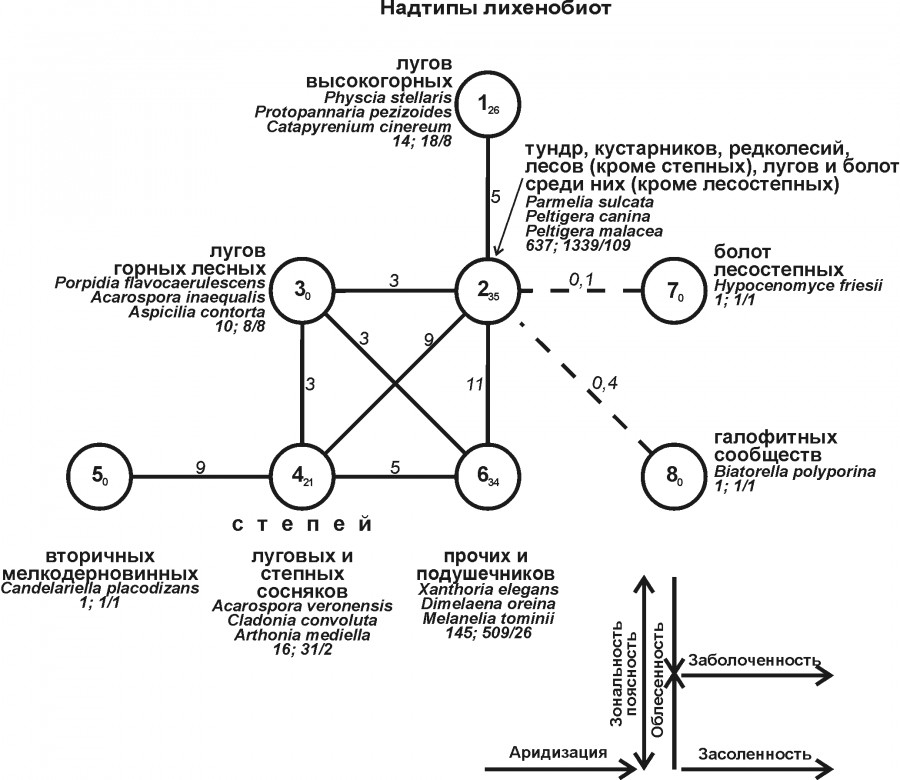
Рис. 2. Структура территориальной неоднородности лихенобиоты Западной Сибири на уровне надтипа. Рядом с номерами надтипов приведены названия занимаемых ими биотопов. Три лидирующих по ценотической значимости вида и суммарное по группе значение ее, а также общее число встреченных видов и через косую черту – фоновых. Фоновыми считали виды, ценотическая значимость которых в среднем по таксону классификации равна или больше единицы. Остальные условные обозначения как на рис. 1
Fig. 2. Structure of the lichenobiotic territorial heterogeneity of Western Siberia at the level of the supra-type. The names of biotopes occupied by supra-types are given next to the numbers of supra-types, Three species with the leading cenotic significance of the species, the total value of the species in the group and the total number of species encountered, and after slash - background ones. Background species were considered those with the cenotic significance, on average, equal to or greater than one in the taxon of classification. The remaining symbols as in Fig. 1
Территориальная изменчивость лихенобиоты в основном совпадает с трендами по зональности в сочетании с поясностью, а также с облесенностью, заболоченностью и засоленностью. Деление на типы сообществ и их характеристики приведены на рис. 3. Пространственные тренды по типам лихенобиоты те же, что и по надтипам.
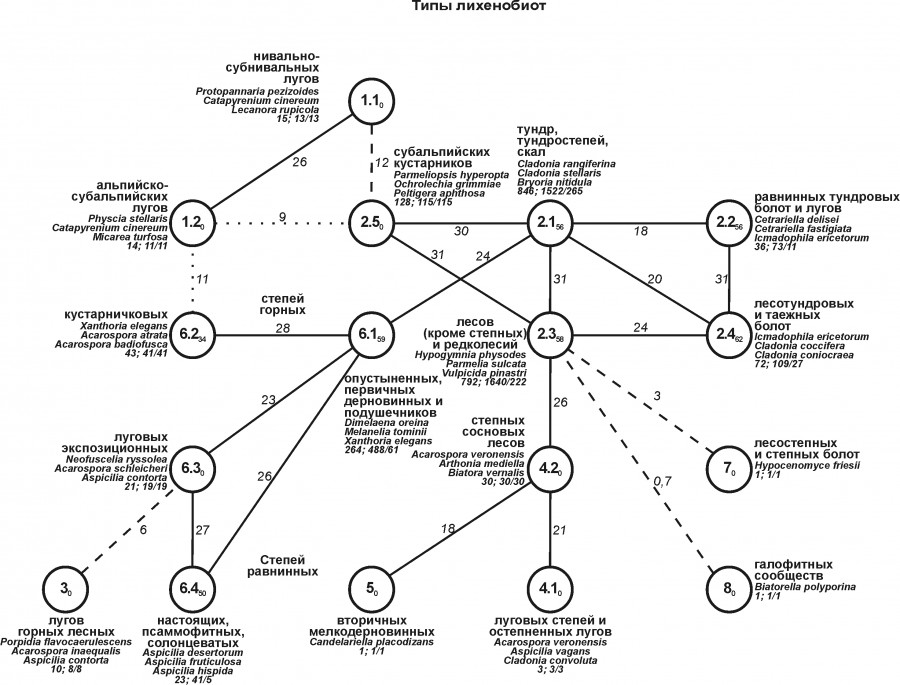
Рис. 3. Структура территориальной неоднородности лихенобиоты Западной Сибири на уровне типа. Условные обозначения как на рис. 1 и 2
Fig. 3. Structure of the lichenobiotic territorial heterogeneity of Western Siberia at the level of the type. Symbols as in Fig. 1 and 2
Типолого-хорологическая неоднородность лихенобиоты
Итоговое представление о пространственной дифференциации лихенобиоты Западной Сибири, сложившееся в результате многомерного анализа данных, их обобщения, классификации, выявления пространственно-типологической структуры и основных трендов, призвана визуально отобразить обзорная карта (рис. 4), составленная в прямой конической равнопромежуточной проекции. При ее подготовке в качестве картографической основы для равнинной части региона взят электронный вариант карты «Растительность Западно-Сибирской равнины» (Ильина и др., 1976) в масштабе 1:1500000, оцифрованной сотрудниками ЦСБС СО РАН, а для горной части использована ландшафтная карта в масштабе 1:1000000 из опубликованной в 2001 г. в виде компакт-диска «Геоинформационной системы Алтае-Саянского экорегиона». Дополнительным источником картографической информации послужила эколого-фитоценотическая карта Южной Сибири в масштабе 1:1000000 (Седельников и др., 2005).
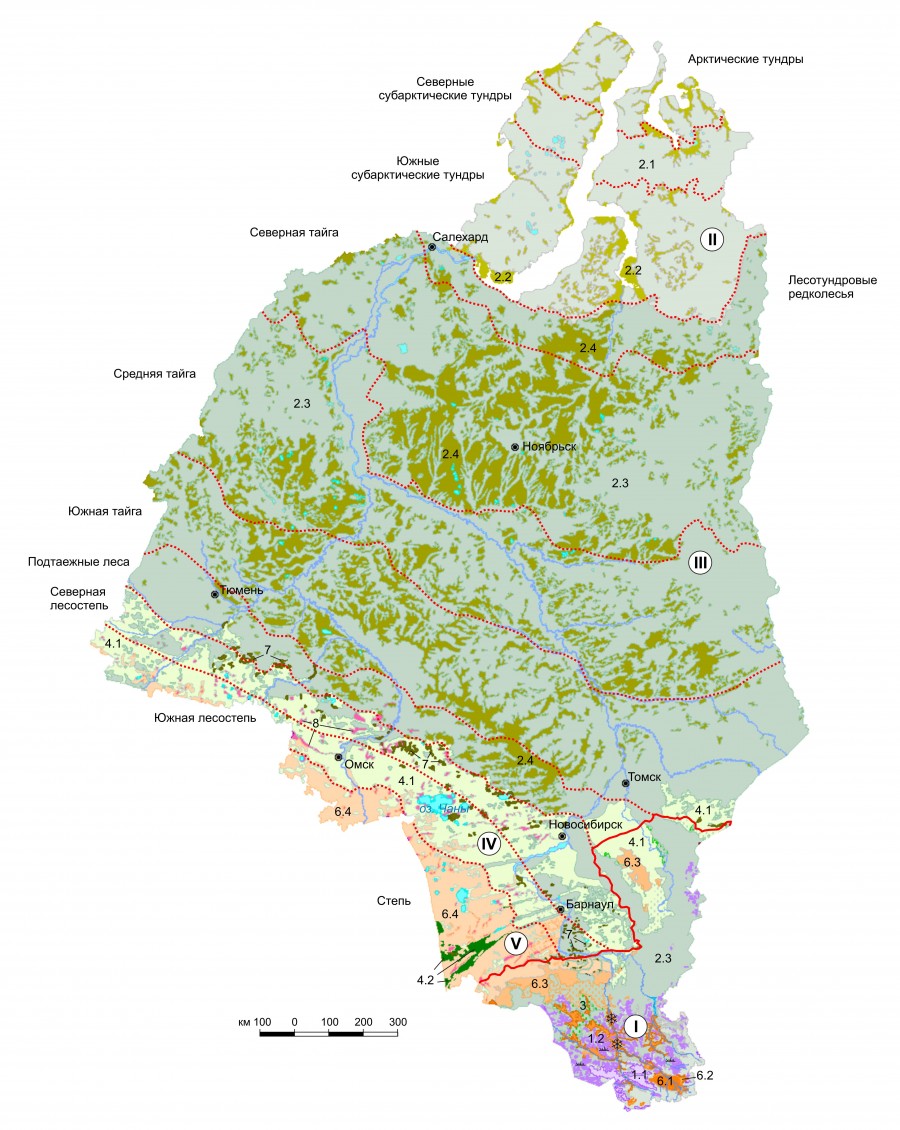
 Рис. 4. Пространственная дифференциация лихенобиоты Западной Сибири (Западно-Сибирская равнина, Алтайская и Кузнецко-Салаирская горные области). Классификацию и характеристики лихенобиоты см. на рис. 2 и 3
Рис. 4. Пространственная дифференциация лихенобиоты Западной Сибири (Западно-Сибирская равнина, Алтайская и Кузнецко-Салаирская горные области). Классификацию и характеристики лихенобиоты см. на рис. 2 и 3
Fig. 4. Spatial differentiation of lichenobiota of Western Siberia (West Siberian Plain, Altai and Kuznetsk-Salair mountain regions). For the classification and characteristics of the biota, see Fig. 2 and 3
Визуальный анализ карты позволяет говорить о наличии пяти типолого-хорологических лихенобиотических полос. Первая из них образована выделами, отражающими распространение лишайников в высокогорных лугах, остальные состоят из трех групп выделов. В первую группу входят преобладающие по площади равнинные выделы, во вторую – их горные аналоги, в третью – равнинные включения из выделов, значительно меньших по площади. Таким образом, лихенобиотические полосы сформированы в основном выделами с наибольшей площадью на равнине, с «выплесками» их в горную часть территории и мелкими включениями, отражающими распространение «дополнительных» таксонов, относящихся к тем же или иным типам либо подтипам. Подтипы каждого типа окрашены в гамме определенного цвета. Следует отметить, что типолого-хорологические полосы, обозначенные на карте римскими цифрами, четких границ не имеют. Они проникают друг в друга, образуя «зоны» пересечения, которые фактически входят в обе граничащие полосы. В легенде карты приведен состав лихенобиот по типам и подтипам, входящим в указанные полосы.
Экологическая организация разнообразия лихенобиот
Анализ классификаций видов и сообществ лишайников свидетельствует о корреляции (по предметным соображениям это, как правило, и причинная зависимость) неоднородности в распространении видов лишайников и изменчивости лихенобиоты с девятью факторами среды. Наиболее значим из них тип растительного покрова (тундровый, лесотундровый, редколесный, кустарниковый, лесной, луговой, степной, болотный и свободный от растительности скальный субстрат, здесь и далее под терминами «скалы» и «скальность» мы понимаем весь набор скалистых биотопов – скал, выходов коренных пород и курумов). Свободные субстраты на стволах, ветвях и почвах входят в соответствующие градации растительности. Тип растительности на региональном уровне определяют широтные или высотные различия по тепло- и влагообеспеченности. Кроме того, с одной стороны, сказывается угнетающее лишайники воздействие конкурентного преимущества высших растений, а с другой – положительное влияние наличия незанятого субстрата на стволах и ветвях. Этим обусловлена высокая степень аппроксимации указанными режимами территориальной неоднородности лихенобиот – 66 % дисперсии матрицы коэффициентов сходства 61 варианта лихенобиот (таблица). Примерно в 1.5 раза меньше влияние теплообеспеченности (в зональном и высотно-поясном аспектах), втрое – облесенности, в 4.5 раза – зональности, взятой отдельно от поясности. Связь с остальными факторами значительно слабее и колеблется в пределах от 2 до 8 % дисперсии. Множественная интегральная оценка вследствие взаимной корреляции между факторами составляет 77 %. Так, оценка влияния облесенности добавляет к аппроксимации первым (наиболее значимым) режимом всего 2 %, а теплообеспеченности – 7 % дисперсии. Значимость остальных факторов полностью входит в совокупную оценку по предыдущим факторам и режимам, поскольку включение их в расчеты не увеличивает снятую дисперсию.
Таблица. Сила и общность связи пространственно-типологической неоднородности биоты лишайников и лихенофильных грибов Западной Сибири и факторов среды (учтенная дисперсия матрицы коэффициентов сходства, %)
Table. Strength and generality of the connection between the spatial-typological heterogeneity of lichen biota and lichenophilic fungi of Western Siberia and environmental factors (the considered variance of the matrix of similarity coefficients, %)
| Фактор, режим | Оценка | |
| индивидуальная | нарастающим итогом | |
| Тип растительного покрова, наличие свободного субстрата | 66 | 66 |
| Облесенность | 33 | 68 |
| Теплообеспеченность (в зональном и высотно-поясном аспектах) | 24 | 75 |
| Зональность | 16 | 75 |
| Скальность | 8 | 75 |
| Поясность | 7 | 75 |
| Заболоченность | 6 | 75 |
| Засоленность | 6 | 75 |
| Рельеф (горы – равнина) | 3 | 75 |
| Классификационные и структурные режимы и факторы совместно | 79 | 90 |
| Множественный коэффициент корреляции | – | 0.95 |
Индивидуальная оценка рассчитана по каждому фактору (режиму) отдельно при условии, что никакие другие факторы неизвестны. Расчет нарастающим итогом выполнен по рассматриваемому фактору и совокупности всех факторов, имеющих более высокие оценки. Это позволяет оценить степень скоррелированности факторов такого набора. В природе, как правило, среди них нет ортогональных, поскольку все факторы в той или иной мере скоррелированы. Поэтому простая сумма оценок связей по всем факторам больше 100 %, чего быть не может. Множественная оценка по всем факторам и режимам с учетом корреляции связей составляет 90 % дисперсии матрицы сходства.
Множественный коэффициент корреляции всех факторов и режимов равен 0.95, то есть информативность изложенных представлений достаточно велика. Указанная оценка не включает в себя представления о типолого-хорологических лихенобиотических полосах, поскольку они выделены не по сходству биот при кластерном анализе территориальной дифференциации лихенобиот, а чисто умозрительно, по характеру изображения на представленной карте, т. е. с учетом хорологической общности.
Обсуждение
Растительность и сообщества животных Западно-Сибирской равнины, Алтайской и Кузнецко-Салаирской горных областей к настоящему времени хорошо изучены. Это позволяет сравнить общий характер территориальной изменчивости лихенобиот, растительности и животного населения. Однако сопоставление биот разных систематических групп осложняют существенные различия в степени полноты собранных данных и их дробности. Так, по лихенобиоте не обследованы пойменные, а также селитебные и агроландшафты. Кроме того, по животному населению, в отличие от лихенобиот, ландшафты описаны раздельно по провинциям Алтайской горной области. Выборка по населению птиц включает водные экосистемы, в то время как по остальным биотическим группам реки и водоемы не обследованы. Поскольку повторное агрегирование стали использовать сравнительно недавно, все сопоставления далее проведены по результатам первого разбиения. Часть классификаций выполнена только по равнинной части Западной Сибири (растительность, гумус почв, подземная фито- и мортмасса, а также население беспозвоночных), поэтому пространственные изменения лихенобиот в этих случаях для сравнимости проанализированы с игнорированием в классификации горных и необследованных сообществ.
По данным за первую половину лета, в условно тундровую группу входят орнитокомплексы только арктической и северной субарктической подзон, а южной субарктической тундры вместе с орнитокомплексами лесотундровой зоны и северной тайги образуют условно лесотундровые сообщества. Лесное население начинается к югу от границы между северной и средней тайгой. По орнитокомплексам гораздо четче, чем по лихенобиотам, отличаются сообщества суходолов и болот, особенно верховых. Население птиц болот не объединено по сходству, как сообщества лишайников, в один тип с биотой горных и равнинных тундр (Равкин и др., 2016).
Пространственные структуры населения птиц второй половины лета и лихенобиоты, в принципе, близки. В самом общем виде суть территориальной изменчивости по этим биотическим группам сводится к соответствию неоднородности растительного покрова, хотя у каждой из групп выявлены специфичные отклонения. Так, по лишайникам горные и равнинные варианты лесов и тундр объединены в один тип лихенобиот, в то время как по населению птиц аналогичная однотипность свойственна только лесным сообществам. По населению птиц Западной Сибири, по данным за вторую половину лета, четко виден контраст между сообществами северной и средней тайги (Равкин и др., 2007). Северотаежные сообщества, так же как в первой половине лета, входят в лесотундровый тип населения вместе с южной субарктической тундрой и лесотундрой, а средне- и южнотаежные – в лесной, включающий и более южные орнитокомплексы – лесостепные, а также горнолесные. По лишайникам подобных различий не выявлено. Кроме того, по лихенобиоте четче видны отличия болотных и незаболоченных сообществ, в то время как по птицам они объединены после усреднения открытых и облесенных болот в единые типы населения с зональными орнитокомплексами. По населению птиц ярче выражены различия сообществ субнивальных и альпийско-субальпийских тундр и лугов. Лихенобиоты последних на уровне типа тоже разделены и отличаются от сообществ горных тундр, входящих в один тип с равнинными аналогами. По птицам это разные типы сообществ. Как по птицам, так и по лихенобиоте сообщества степей в общем делятся на луговые (субгумидные) и настоящие (сухие), а в горах – еще и криофитно-аридные.
По мелким млекопитающим в общих чертах прослежены те же изменения, что и по птицам, но без четких различий между населением северо- и среднетаежных местообитаний и несколько большими отличиями населения болот и незаболоченных территорий (Равкин и др., 2009). Это сближает характер пространственной изменчивости сообществ мелких млекопитающих и лихенобиот.
Наиболее велики отличия от ранее рассмотренных биотических групп у наземных холоднокровных позвоночных – земноводных и пресмыкающихся (Равкин и др., 2008). По этим группам на фоне зональной специфики доминирует влияние отдельных лимитирующих факторов среды. В их список входят теплообеспеченность, увлажнение и кормность. По другим группам позвоночных из-за большего числа видов и их экологической разнородности аналогичные изменения плавны и в целом больше совпадают с зонально-подзональным делением. По холоднокровным таксоны классификации дифференцированы по занимаемым биотопам: экстремальным, субэкстремальным, пессимальным (отдельно равнинным и горным), а также субоптимальным и оптимальным (только для земноводных). Лимитировать распределение этих позвоночных может один из перечисленных факторов независимо от благополучного состояния по остальным. Это, как правило, и порождает включение в один таксон сообществ, совершенно различных по зональности, подзональности, заболоченности и типу растительности. При этом степень совпадения границ таксонов по рассматриваемым биотам выше в тундрово-лесотундровой полосе и меньше – в более южных частях региона.
По Западно-Сибирской равнине при сравнимых подходах и принципах классификации все фитоценозы разделены на четыре типа: 1 – тундровые; 2 – лесотундрово-северотаежные; 3 – лесные (от средней тайги до лесостепи, включая болотные сообщества) и 4 – степные (Пространственно-типологическая дифференциация…, 2011). В общих чертах классификация экосистем, составленная авторами этой публикации, по биомассе пяти жизненных форм растений на уровне типа совпадает с таковой по лихенобиотам и населению птиц. Более общий вид имеет классификация экосистем по гумусу почв, морт- и подземной фитомассе. В ней выделены только три типа экосистем: 1 – незаболоченных территорий; 2 – болот тундровой зоны и 3 – болот бореальных и суббореальных. Очевидно, что неоднородность в данном случае может быть объяснена различиями по заболоченности и макрозональности в виде деления на тундровую и всю остальную более южную часть. Эта классификация генерализована в большей степени, чем по лихенобиоте и остальным рассмотренным группам.
По беспозвоночным Западно-Сибирской равнины выделено четыре типа экосистем: 1 – тундровой зоны; 2 – лесотундрово-северотаежных и пойм средней тайги; 3 – лесов и болот в пределах от средней тайги до лесостепи и 4 – прочих степных и лесостепных экосистем и пойм южной тайги. Таким образом, деление равнинной территории Западной Сибири по населению беспозвоночных ближе всего к таковому по биомассе пяти жизненных форм растений, а по лихенобиотам и орнитокомплексам на уровне первого разбиения значимо отличается по болотным и незаболоченным экосистемам. По наземным позвоночным классификация имеет большую степень обобщения. Первое разбиение соответствует трем типам сообществ: 1 – тундровых; 2 – лесотундрово-северотаежных и 3 – таежно-степных (от средней тайги до степной зоны включительно). Таким образом, четкого разделения на лесные и степные сообщества (если считать лесостепь подзоной степной зоны), как по наземным беспозвоночным, для данной группы не выявлено. Бóльшая обобщенность по позвоночным в сравнении с лихенобиотой обусловлена отсутствием значимых отличий болотных сообществ и меньшей дифференциацией в таежно-степной части региона.
Классификация экосистем, выполненная на матрице средних коэффициентов сходства по всем четырем биотическим группам, существенно отличается от таковой по лихенобиотам. Семь из восьми типов экосистем входят в три основных типа лихенобиот. Лишь единственная группа итоговой классификации – лесостепная с проникновением в подтаежные леса и степную зону – представлена, хотя и в усеченном виде, двумя аналогичными типами по лишайникам. Резюмируя, можно отметить, что классификация равнинных экосистем Западной Сибири в значительной мере соответствует общепринятым представлениям о зональных подразделениях этого региона, хотя и с делением на незаболоченные территории, болота, луга и с целым рядом небольших отклонений. Первое из этих отклонений сводится к объединению экосистем лесотундры и северной тайги. Второе проявляется в выделении в качестве Лесного типа лесных экосистем только средней и южной тайги. Третье – в объединении в один тип биоценозов подтаежных лесов, лесостепи и степной зоны. По лихенобиоте все незаболоченные и заболоченные, лесные и открытые территории на всем пространстве от тундр до южных границ лесостепи, за редким исключением, объединены в один основной надтип сообществ. Однако на уровне типа сходство гораздо выше. Таким образом, имеет место отмеченная ранее подвижность границ (Бунге, 1967) в зависимости от степени агрегации данных, которую определяет специалист-аналитик, хотя классификации, конечно, отражают и внутренний уровень сходства-различия биотических групп.
По равнинной и горной частям Западной Сибири на составленных картах больше всего типолого-хорологических полос прослежено по населению птиц (7). По сравнению с лихенобиотой эти сообщества не образуют единой полосы по равнинным и горным тундрам, хотя территории лесотундровой и лесной полос объединены, а высокогорные луга представлены отдельной полосой. По мелким млекопитающим всего на одну полосу больше, чем по лихенобиотам, и основными из них остаются только Тундровая, Лесотундрово-северотаежная, Лесная (от средней тайги до лесов лесостепной зоны) и две Степных (луговых и настоящих степей). По земноводным и пресмыкающимся число полос такое же, как по лихенобиотам, хотя принципы их формирования и границы не полностью совпадают. Так, пессимальные для холоднокровных животных территории различны в горах и на равнине, а субоптимальные и оптимальные занимают общую часть пространства, занятого сходными лихенобиотами.
Количество типолого-хорологических полос в пределах Западно-Сибирской равнины по исследованным биотопам колеблется в пределах от трех до четырех. Обычно это Тундровая, Лесотундрово-северотаежная, Лесная (от средней тайги до подтаежных лесов) и Лесостепная. Отклонения сводятся чаще всего к делению лесостепной и степной частей территории. Они объединены, как на карте растительности (Ильина и др., 1976), а по беспозвоночным, так же как по экосистемам в целом, включают, кроме того, и сообщества подтаежных лесов. По биомассе основных групп растений лесостепная полоса объединена с лесной (начиная со средней тайги). По лишайникам лесные выделы в лесостепи отнесены к Редколесно-лесной полосе, а степные разделены на сообщества луговых и настоящих степей. По позвоночным вся территория от средней тайги до степной зоны образует единую типолого-хорологическую полосу. Принципиально отличается отображение неоднородности по гумусу почв, морт- и подземной фитомассе. По неоднородности этих компонентов биоты доминирует влияние увлажнения, а зональность представлена лишь на втором шаге деления.
Таким образом, несмотря на то, что ведущим фактором пространственной дифференциации лихенобиот, растительности и населения животных выступает неравномерность распределения тепла и влаги, вследствие различий в толерантности лишайников, растений и животных, границы и состав типолого-хорологических полос существенно отличаются. При этом авторы настоящего сообщения вовсе не считают необходимым изменять зональные и подзональные границы, проведенные умозрительно, по облику растительности. Эти границы идеализированы (концептуализированы), они теоретические по своей сути. Мы лишь обращаем внимание читателей на несовпадение их на исследованной территории при однотипном формализованном анализе по группам организмов, для которых имеется соответствующая информация. Подробнее об этом см. в статье Ю. С. Равкина, И. Н. Богомоловой и С. М. Цыбулина (2017).
Заключение
Статья, посвященная пространственно-типологической дифференциации лихенобиоты Западной Сибири, продолжает серию сходных сообщений, ссылки на которые приведены в тексте. Неоднородность распределения лишайников и лихенофильных грибов может быть отображена тремя надтипами распространения: Тундровым, Лесным и Степным. Дальнейшее деление на типы отражает различия во встречаемости отдельных видов в экосистемах равнинных тундр, болот, лесотундровых комплексов, тундростепей, скальных обнажений, горных и равнинных лесов и степей. По сходству сообществ лишайников и лихенофильных грибов выделено восемь надтипов: 1 – горных нивально-субнивальных и альпийско-субальпийских лугов; 2 – тундр, кустарников, редколесий и лесов (исключая степные), болот (кроме лесостепных и степных), а также лугов в пределах всех перечисленных экосистем (не считая сообщества лишайников, которые могут быть выделены в отдельный таксон надтипового ранга). Остальные надтипы включают лихенобиоты степей и сосновых лесов степной зоны, а также лесостепных и степных болот и галофитных сообществ. Указанные надтипы могут быть разделены на 17 типов лихенобиот, в основном по различию сообществ нивально-субнивальных и альпийско-субальпийских лугов, тундр, тундростепей, болот, лесов и редколесий.
Иерархическая классификация лихенобиот в картографическом отображении демонстрирует наличие пяти типолого-хорологических полос. Одна из них ограничена распространением нивально-субнивальных и альпийско-субальпийских лугов и сильно фрагментирована, а остальные простираются на равнинных территориях с «выплесками» в горы и включением меньших по площади «инородных» выделов на равнине. Вторая полоса составлена биотами тундр, третья – лесов и редколесий, четвертая и пятая – степей, соответственно луговых и настоящих.
Информативность представленных классификаций вполне достаточна для прогнозирования распределения видов лишайников и лихенофильных грибов и неоднородности лихенобиот на необследованных участках Западной Сибири. Степень обобщенности представлений о дифференциации изученных биотических групп по сходству видового состава и встречаемости, а также неоднородность территорий по гумусу почв, морт- и подземной фитомассе как производных биоты существенно различается, причем не только в силу их специфичности, но и по причине дробности характеризующих признаков. Так, наиболее простой структурой на уровне типа (по результатам первой агрегации) можно считать классификацию экосистем по биотическим производным, растительности и населению наземных позвоночных (по 3 типа). В 1.5 раза большая дробность свойственна сообществам наземных беспозвоночных, земноводных и пресмыкающихся, а также мелких млекопитающих (5 типов) и в 2–2.5 раза – по орнитокомплексам и экосистемам в целом по указанным биотическим группам и их производным. Максимальное разнообразие по этому показателю свойственно лихенобиотам (17 типов). Типологическое разнообразие в некоторой степени зависит от числа признаков (видов или групп), по которым проведена классификация, хотя и не строго пропорционально. Так, при пяти признаках по гумусу почв выделено три типа, а по другим группам растений и животных, при числе признаков от шести до 44, – по пять типов, при количестве признаков от 291 до 369 – шесть – восемь типов и 17 типов – при 1942 признаках (формах лишайников и лихенофильных грибов). Исключение представляют только позвоночные животные. По ним при 353 видах после первой агрегации выделено всего три типа.
Таким образом, отличия в границах таксонов классификаций разных сообществ зависят от их интегральной средовой толерантности, а число выделяемых типов сообществ – в некоторой степени и от количества видов растений и животных, если эти признаки для классификации не сведены к групповым представлениям.
Библиография
Биологический энциклопедический словарь / Под ред. М. С. Гилярова. М.: Советская энциклопедия, 1986. С. 595.
Бунге В. Теоретическая география . М.: Прогресс, 1967. 279 с.
Ильина И. С., Лапшина Е. Н., Лавренко Н. Н., Мельцер Л. И., Романова Е. А., Богоявленский Б. А., Махно В. Д. Растительность Западно-Сибирской равнины. Карта масштаба 1:1500000 . М.: МГУГК, 1976.
Западная Сибирь / Под ред. Г. Д. Рихтера. М.: Изд-во АН СССР, 1963. 488 с.
Куперштох В. Л., Миркин Б. Г., Трофимов В. А. Сумма внутренних связей как показатель качества классификации // Автоматика и телемеханика. 1976. № 3. С. 133–144.
Куперштох В. Л., Трофимов В. А. Автоматическое выявление макроструктуры системы // Проблемы анализа дискретной информации. Новосибирск, 1975. Ч. 1. С. 67–83.
Миркин Б. Г. Анализ качественных признаков и структур // Статистика. 1980. 319 с.
Пространственно-типологическая дифференциация экосистем Западно-Сибирской равнины . Сообщение I–V // Сибирский экологический журнал. 2011. Т. 18. № 3–6.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М. Экологическая организация флоро-фаунистической неоднородности Северной Евразии // Принципы экологии. 2017. № 1. С. 104–116.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М., Железнова Т. К., Торопов К. В., Вартапетов Л. Г., Миловидов С. П., Юдкин В. А., Жуков В. С., Гуреев С. П., Покровская И. В., Касыбеков Э. Ш., Ананин А. А., Бочкарева Е. Н. Пространственно-типологическая неоднородность и экологическая организация летнего населения птиц Срединного региона Северной Евразии // Сибирский экологический журнал. 2016. Т. 9. № 1. С. 103–116.
Равкин Ю. С., Богомолова И. Н., Цыбулин С. М., Жуков В. С., Ливанов С. Г. Пространственно-типологическая классификация населения мелких млекопитающих Западной Сибири // Вестник Новосибирского государственного университета. Серия: Биология, клиническая медицина. 2009. Т. 7. Вып. 4. С. 86–94.
Равкин Ю. С., Куперштох В. Л., Трофимов В. А. Пространственная организация населения птиц // Равкина Ю. С. Птицы лесной зоны Приобья. Новосибирск: Наука, 1978. С. 253–269.
Равкин Ю. С., Ливанов С. Г. Факторная зоогеография . Новосибирск: Наука, 2008. 205 с.
Равкин Ю. С., Цыбулин С. М., Вартапетов Л. Г., Ливанов С. Г., Торопов К. В., Жуков В. С., Юдкин В. А. Пространственная структура и организация населения птиц Алтая и Западно-Сибирской равнины // Сибирский экологический журнал. 2007. Т. 14. № 6. С. 877–884.
Равкин Ю. С., Цыбулин С. М., Ливанов С. Г., Торопов К. В., Куранова В. Н., Стариков В. П., Чеснокова С. В. Картографический анализ населения земноводных, пресмыкающихся и птиц Западно-Сибирской равнины и Алтая // Сибирский экологический журнал. 2008. Т. 15. № 5. С. 745–750.
Реймерс Н. Ф. Азбука природы (микроэнциклопедия биосферы) . М.: Знание, 1980. 208 с.
Реймерс Н. Ф. Природопользование (словарь-справочник) . М.: Мысль, 1990. 640 с.
Седельников В. П., Лапшина Е. И., Королюк А. Ю., Валуцкий В. И., Ермаков Н. Б., Ершова Э. А., Макунина Н. И., Мальцева Т. В. Среднемасштабное картирование растительности гор Южной Сибири // Сибирский экологический журнал. 2005. Т. 12. № 6. С. 939–953.
Седельникова Н. В. Видовое разнообразие лихенобиоты Западной Сибири и оценка участия видов лишайников в основных ее горных и равнинных фитоценозах . Новосибирск: Академическое изд-во «ГЕО», 2017. 612 с.
Сержантов В. Ф. Введение в методологию современной биологии . Л.: Наука, 1972. 282 с.
Благодарности
Исследования, послужившие основой для настоящей публикации, выполнены в рамках программы ФНИ государственных академий на 2013–2020 гг. и СО РАН (проекты № ААА-А-17-117012610055–3.0312-2016-0005 и АААА-А16-116121410122-4) и частично по «Программе повышения конкурентоспособности ТГУ».

 © 2011 - 2024
© 2011 - 2024