



Введение
Цикличность в динамике численности лесных полевок изучается достаточно длительное время (Европейская рыжая полевка, 1981). Описана правильная цикличность в динамике численности этих полевок в северных популяциях (Stenseth, 1985). Особенно интенсивно изучали динамику лесных полевок в Фенноскандии, как в связи с воздействием хищников, так и с влиянием кормового фактора (Hansson, Stenseth, 1988; Hanski et al., 1993; Turchin, Hanski, 1997). Ряд работ посвящен изучению цикличности лесных полевок, причем в них даются не просто описания динамики и установление циклов по среднему расстоянию между пиками численности, но используются и спектральные оценки для выявления параметров циклов (Максимов, Ердаков, 1985; Бобрецов и др., 2000; Жигальский, Кшнясев, 2000; Чернявский, Лазуткин, 2004; Ердаков, 2011).
В Японии цикличность динамики рыжих лесных полевок изучали на о-ве Хоккайдо, исследовались спектры, рассчитанные по многолетним наблюдениям по всей периферии острова (Bjørnstad et al., 1996; Stenseth et al., 1996), анализировались закономерности динамики красно-серой полевки в северной части Хоккайдо и рассматривался вклад многолетней цикличности в сезонные и географические адаптации популяций (Saitoh et al., 1999).
В Северной Америке многолетние материалы по изучению динамики численности красной полевки также рассматривались с точки зрения цикличности. Полученные данные сравнивали с цикличностью в популяциях красной полевки, опубликованной для Фенноскандии (Hansson, Henttonen, 1985; Henttonen et al., 1985). Там же продолжается полемика с B. H. McArdle и K. J. Gaston (1992) об определении циклических и нециклических популяций. Авторы используют для такого определения индекс S (Boonstra, Krebs, 2012). Однако ранее нами уже было выяснено, что на разных участках временного ряда (отрезки из 4–5 значений) индекс цикличности Левонтина (S) заметно варьирует, определяя одну и ту же популяцию то как «цикличную», то как «нецикличную» (Ердаков и др., 1990). Кроме того, на американских популяциях проверялась зависимость цикличности от различных факторов: кормовой базы, конкуренции, цикличности климата. Рассматривая ранее цикличность динамики рыжих лесных полевок в Западной Сибири, мы отмечали, что сложность адаптации этих видов грызунов определяется, в частности, и приспособлением к одним и тем же жизненно важным для них внешним синхронизаторам (из-за значительной эколого-физиологической близости видов). Они вынуждены разделяться во времени, чтобы снизить напряженность конкурентных отношений. Разделение популяций достигается за счет варьирования значений фаз, периодов и мощностей циклов динамики численности (Ердаков, Панов, 1987).
Несмотря на обширность уже проведенного изучения цикличности динамики численности красной полевки, многие участки ее ареала еще не затронуты такими исследованиями. Это затрудняет создание интегральной картины циклов ее динамики численности на всем ареале. Наша работа дополняет уже имеющиеся сведения о циклах, расширяя возможности сравнения. Целью исследования является описание цикличности динамики численности красной полевки на значительной территории ее ареала при унификации сведений.
Задачи сводились к построению спектров ритмов многолетней динамики численности в разных регионах ареала; определению параметров ритмов; выявлению особенностей цикличности и возможных географических закономерностей ее изменений; описанию инвариантных частей спектра ритмов численности; поиску вероятных ритмоводителей для многолетней численности циклов красной полевки.
Материалы
Основу материала составили опубликованные данные (Ердаков, 2011; Ердаков, Панов, 1987; Ердаков и др., 1990), собранные на стационарах лаборатории экологии Биологического института СО АН СССР (ныне ИСиЭЖ СО РАН).
Кроме того, для описания многолетней цикличности в различных регионах использованы опубликованные данные литературных источников со следующих территорий: Карелия (Ивантер, Моисеева, 2015); Центрально-Лесной биосферный заповедник (Истомин, 2009); Печоро-Илычский биосферный заповедник (Бобрецов и др., 2000); Баргузинский биосферный заповедник (собственные данные); Магаданский заповедник, среднее течение р. Челомджи (Чернявский, Лазуткин, 2004); Аляска, Юкон (West, 1982); Канада, северо-западные территории (Fuller, 1977); окрестности Новосибирска 1980–2014 гг. (неопубликованные данные В. В. Панова).
Методы
Для выявления скрытых колебаний в численности применяли быстрый анализ Фурье. Эмпирически определяемые параметры счета: шаг суммирования, длина автокорреляционной функции, форма и ширина корреляционного окна. Вычисления проведены стандартно (Ердаков, 2011). Использованы счетные программы, находящиеся в собственности ИСиЭЖ СО РАН. Оценка спектральной плотности мощности проводилась методом Уэлча (Welch) (Марпл, 1990).
Для трендов проверяли гипотезу об отличии коэффициентов от нуля и рассчитывали коэффициент детерминации R2. Полученные значения отражаются в тексте только в том случае, когда нулевая гипотеза о равенстве обоих коэффициентов регрессии нулю отбрасывается. В противном случае линейная регрессия исключается из модели процесса.
Для ограничения числа гармоник в модели использовали тест Льюнга – Бокса для автокорреляции остатков с заданным критерием (http://www.machinelearning.ru/wiki/index.php?title). Тест проверяет остатки на некоррелированность. Это означает, что после включения в модель первой самой мощной гармоники автокорреляция остатка не вышла за доверительные границы.
Обработка данных проведена методами спектрального анализа, для статистической обработки использована программа Past 3.17.
Результаты
Сравнение статистических показателей средней численности и ее изменений показывает значительные различия в разных регионах, хотя везде численность этого вида отслеживалась в его обычных лесных местообитаниях. Наиболее низкая численность зафиксирована для Карелии. Здесь учеты велись по всей территории (Ивантер, Моисеева, 2015), и эти наблюдения самые продолжительные. Тем не менее численность здесь самая низкая и к тому же подвержена наиболее сильным колебаниям (табл. 1).
Невелика численность красной полевки и на территории Печоро-Илычского заповедника, самой северной территории из представленного ареала. Как и в Карелии, она сильно варьирует и имеет отрицательный тренд за время наблюдений. Несколько выше численность красной полевки на юге Западной Сибири (Академгородок, Новосибирск). Красная полевка здесь является доминирующим видом наряду с красно-серой полевкой и лесной мышовкой. Численность ее весьма стабильна, среди исследуемых данных она варьирует менее других и имеет тенденцию к медленному повышению (см. табл. 1).
Таблица 1. Статистические показатели численности в популяциях красной полевки
| № | Регион | n | Численность M ± m | σ | CV | тренд |
| 1 | Карелия | 55 | 0.03 ± 0.008 | 0.057 | 190 | 0.075 – 0.002 * t; R2 = 0.198; α = 0.01 |
| 2 | Центрально-Лесной заповедник | 25 | 4.93 ± 1.117 | 5.583 | 113.25 | -0.610 + 0.461 * t |
| 3 | Печоро-Илычский заповедник | 45 | 1.59 ± 0.321 | 2.154 | 135.47 | 3.026 – 0.065 * t; R2 = 0.157; α=0.01 |
| 4 | Бараба, Ровенское | 24 | 1.34 ± 0.21 | 1.01 | 75.37 | 1.612 – 0.024 * t |
| 5 | Новосибирск, Академгородок | 35 | 2.21 ± 0.170 | 1.007 | 45.57 | 1.746 + 0.027 * t |
| 6 | Баргузинский заповедник | 55 | 7.85 ± 0.568 | 4.213 | 53.67 | 7.807 + 0.002 * t |
| 7 | Магадан | 20 | 24.43 ± 2.793 | 12.8 | 52.39 | 13.262 + 1.117 * t |
| 8 | Канада, Юкон | 19 | 6.87 ± 1.733 | 7.553 | 109.94 | 3.628 + 0.360 * t |
| 9 | Канада, NWT | 19 | 4.80 ± 1.15 | 5.017 | 104.52 | 8.439 – 0.404 * t |
Примечание. n – число рассматриваемых лет, M – среднее, m – ошибка среднего, σ – квадратичное отклонение, CV – коэффициент вариации, R2 – коэффициент детерминации.
Высокая численность отмечена на Аляске и на территории Центрально-Лесного заповедника. И средние многолетние значения численности, и варьирование численности находятся в близких пределах. Численность в этих местах постепенно нарастает, причем тренд у нее заметный.
Величина тренда может быть заметной, однако он может оказаться недостоверным. При проверке трендов с помощью коэффициента детерминации (R2) выяснилось, что только в двух географических популяциях имеется высоко достоверный отрицательный тренд. Речь идет о северных таежных районах Карелии и Печоро-Илычского заповедника (табл. 1).
Самая высокая численность красной полевки из исследуемых территорий наблюдается в Магаданском заповеднике, где показатели численности многократно превышают даже многолетнюю численность в популяции на р. Юкон (см. табл. 1). При этом варьирование ее невелико, и наблюдается постепенное повышение ее значений за время наблюдений (Чернявский, Лазуткин, 2004).
Итак, численность во всех регионах, из которых мы имеем данные той или иной продолжительности наблюдений, заметно колеблется. В некоторых популяциях такие колебания особенно сильны. Следующим шагом в наших исследованиях будет оценка наборов гармонических составляющих в таких колебаниях численности, то есть их спектры (рис. 1, 2).
Самый длинный ряд наблюдений, который мы анализировали на присутствие циклов, это популяции красной полевки на территории Карелии (Ивантер, Моисеева, 2015). Наибольшей мощностью на нем обладают 5–6-летние гармонические составляющие, а также имеется низкочастотный, примерно 30-летний, цикл (см. рис. 1а). Эта многолетняя цикличность на нашем спектре имеет очень широкое основание, так что при увеличении длины ряда наблюдений в этой полосе частот может оказаться два или несколько пиков. Имеются на спектре из Карелии и множество небольших по мощности гармонических составляющих (примерно 3- и 2-летних).
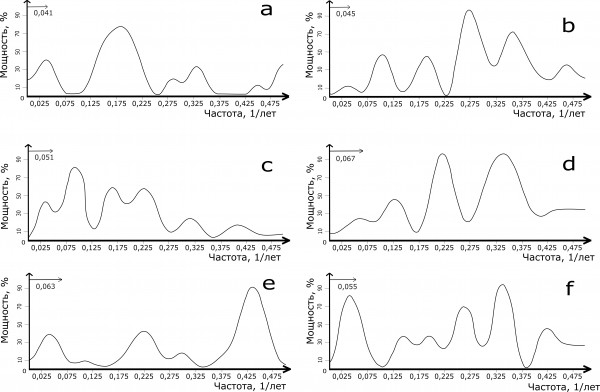
Рис. 1. Спектры цикличности динамики численности красной полевки в разных регионах с запада на восток (a – Карелия, b – Тверь, c – Печоро-Илычский заповедник, d – Новосибирский Академгородок, e – Баргузинский заповедник, f – Магаданский заповедник)
Fig. 1. Spectral densities of the northern red-backed vole in different parts of Russia from West to East (a – Karelia, b – Tver', c – Pechora-Ilych Nature Reserve, d – Akademgorodok in Novosibirsk, e – Barguzin Nature Reserve, f – Magadan Nature Reserve)
Близкий к Карелии спектр ритмов многолетней динамики оказался у популяции красной полевки с р. Юкон (Аляска). Здесь на спектре также доминирует по мощности примерно 5-летний цикл, имеются мощный примерно 3-летний и заметный 20-летний ритмы (см. рис. 2a). Пик этого ритма имеет широкое основание, то есть его параметры не очень точны, повысить ее может удлинение ряда данных, но на Юконе работали с короткими рядами наблюдений. Анализируя свои данные, Boonstra и Krebs (2012) пришли к выводу, что красная полевка в своей динамике здесь показывает 3- и 5-летнюю цикличность. Близкие циклы динамики красной полевки они нашли в Манитобе и Миннесоте (4–5-летние), а также в Пенсильвании (примерно 4-летний цикл). Все это вычислялось по долгосрочным исследованиям, примерно 12–14-летним, что, по нашему мнению, недостаточно для уверенного выявления гармонических составляющих в широком диапазоне частот.
На территории Печоро-Илычского заповедника и в районе Новосибирского Академгородка на протяжении всех лет наблюдений фиксировалась невысокая численность красной полевки. Различия между этими популяциями состояли в том, что вариабельность многолетних значений численности в заповеднике вдвое превосходила таковую в Академгородке (см. табл. 1). В обоих регионах красная полевка доминирует среди грызунов. Периодические составляющие на спектрах отмечены примерно в одних полосах частот, визуальное различие изображений происходит от несовпадения мощностей одинаковых гармоник. Так, приблизительно 3-летняя цикличность доминирует на спектре из Академгородка (см. рис. 1d), на спектре заповедника она также присутствует, но невелика по величине (см. рис. 1c). Мощной цикличностью на спектре и в Новосибирском Академгородке, и в заповеднике отличаются примерно 5- и 3-летние пики. Различие заключается в том, что на спектре динамики красной полевки из заповедника имеется мощный пик в самой низкочастотной области – примерно 30-летний. Сибирский же спектр такой гармонической составляющей не содержит. При этом длина наблюдений в обоих регионах сопоставима по продолжительности.
Наиболее значительная средняя многолетняя численность красной полевки отмечена в Баргузинском и Магаданском заповедниках (см. табл. 1), где этот вид является фоновым, а часто и доминирует. На спектрах цикличности обоих заповедников выделяются доминирующий у красных полевок примерно 3-летний цикл динамики и второй по мощности – более чем 20-летний (см. рис. 1e, f). Анализируя многолетнюю динамику численности красной полевки в Магаданском заповеднике, Ф. Б. Чернявский и А. Н. Лазуткин (2004) также оценивали спектр колебаний ее численности. Доминировал на спектре 3-летний цикл, вторым по мощности оказался более чем 4-летний, а кроме того, был проявлен еще и приблизительно 2-летний. К сожалению, эти авторы не привели параметры рассчитанных колебаний, по изображениям спектров они оцениваются только приблизительно.
Далее, на Аляске и в Канаде цикличность хода численности в популяциях красной полевки меняется. Boonstra и Krebs (2012), применяя спектральный анализ, обнаружили только 3- и 5-летний циклы динамики численности. Мы построили спектры цикличности численности красных полевок из Юкона (West, 1982) и северо-западных территорий (Fuller, 1977) и сравнили их (рис. 2). На спектрах имеются средние по мощности 3- и 4-летние циклы, однако доминируют на них ритмы численности в средних частотах (5–6-летние). Использованные для расчета ряды данных были невелики, поэтому иллюстрирующие их пики имели широкое основание, то есть можно говорить только о полосе частот. У обеих популяций имеются 20-летние колебания численности.
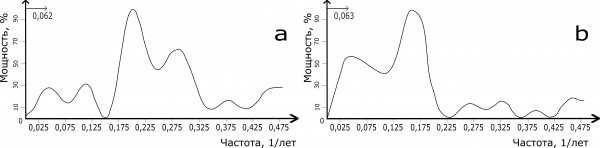
Рис. 2. Спектры цикличности динамики численности красной полевки в Канаде (a – Юкон, b – северо-западные территории)
Fig. 2. Spectral densities of the northern red-backed vole in Canada (a – Yukon, b – Northwest Territories)
Визуальные изображения позволяют примерно оценить соотношения гармонических составляющих по внешнему подобию спектров. Для уверенного сравнения популяционной многолетней цикличности в разных регионах нужны данные расчетов параметров этих спектров, хотя бы величины и мощности каждой имеющейся на них периодичности. Используем для такого сравнения данные из табл. 2.
Наиболее низкочастотные циклы у этих популяций отмечены в Центрально-Лесном и Печоро-Илычском заповедниках, это почти 30-летние циклы. На спектрах они имеют пики с широким основанием, то есть частота определяется недостаточно четко, но синусоида с таким периодом помещается в срок проведенных наблюдений, поэтому такую цикличность фиксирует изображение спектра. Следующая полоса низких частот, характерная для динамики полевки, – примерно 20-летние. Они проявлены на северо-востоке ареала красной полевки – в Баргузинском и Магаданском заповедниках, на Юконе (Аляска) и в Канаде. В Центральной Барабе (Западная Сибирь) и под Новосибирском отмечены 16- и почти 14-летние гармоники, которые имеют заметную мощность (см. табл. 2).
Таблица 2. Соотношение величины и мощности периодических составляющих многолетней динамики в популяциях красной полевки
| Период, лет | 25–35 | 15–20 | 10–14 | 7–9.5 | 4.5–6 | 3.5–4.4 | 2.8–3.3 | 2.0–2.7 | |
| Карелия | 25.6 | – | – | – | 5.4 | 3.5 | 3.0 | 2.5 | 2.2 |
| 0.01 | 0.02 | 0.008 | 0.01 | 0.003 | 0.006 | ||||
| Центрально-Лесной заповедник | 26.3 | – | – | 9.5 | 5.2 | 3.7 | – | 2.8 | 2.1 |
| 0.53 | 1.11 | 1.08 | 1.59 | 1.37 | 0.95 | ||||
| Печоро-Илычский заповедник | 28.4 | – | 10.8 | – | 5.9 | 4.4 | 3.1 | 2.4 | – |
| 0.46 | 0.64 | 0.54 | 0.54 | 0.46 | 0.28 | ||||
| Новосибирский Академгородок | – | 16.3 | – | 7.7 | 4.5 | – | – | 2.9 | 2.2 |
| 0.13 | 0.18 | 0.26 | 0.26 | 0.16 | |||||
| Ровенское | – | 13.8 | – | – | 5.5 | – | 2.4 | – | – |
| 0.40 | 0.38 | 0.24 | |||||||
| Баргузинский заповедник | 25 | – | – | 9.1 | 4.5 | 3.4 | – | 2.3 | – |
| 0.52 | 0.25 | 0.54 | 0.35 | 0.79 | |||||
| Магаданский заповедник | 23.3 | – | – | 6.8 | 5.0 | 3.8 | 2.9 | 2.3 | – |
| 3.87 | 2.55 | 2.56 | 3.56 | 4.15 | 2.86 | ||||
| Канада, р. Юкон | 20.9 | – | – | – | 4.8 | 3.4 | 2.5 | – | 2.0 |
| 1.70 | 3.13 | 2.51 | 1.32 | 1.71 | |||||
| Канада, оз. Сердце | 20.9 | – | – | 6.0 | – | 3.6 | 2.9 | 2.5 | 2.1 |
| 1.63 | 2.12 | 0.81 | 0.93 | 0.52 | 0.93 | ||||
Примечание. Верхняя цифра – период, лет; нижняя – мощность (амплитуда).
Что касается цикличностей численности в остальных полосах, то они характерны практически для всех популяций красной полевки и отмечались многими авторами, которые исследовали именно цикличность в динамике численности этого вида. Гармонические составляющие, выделенные в динамике американских популяций красной полевки, в целом укладываются в своих ритмах в характерную для этого вида картину (табл. 2).
Популяционная цикличность эндогенна, но любые эндогенные ритмы обычно носят затухающий характер. Для формирования устойчивости такие колебания в популяции синхронизируются (подстраиваются) к соответствующим по периоду каким-либо внешним циклам. Такими циклами могут быть, например, климатические изменения, цикличность их обладает значительным постоянством. Не исключено, что подстройка популяционного ритма может происходить к близкому по значению циклу биологически важного для данного вида ресурса (например, пищевому).
Чтобы проверить такую возможность, необходимо выяснить спектральные характеристики климатического фона и ресурсов данного организма. Такая возможность представилась, потому что наряду с многолетними данными динамики численности красной полевки в Печоро-Илычском заповедники были опубликованы сведения об изменениях климата (осадки, ход температур), а также многолетняя динамика плодоношения хвойных видов деревьев (Бобрецов и др., 2000).
Сравнивая цикличность динамики численности красной полевки в Печоро-Илычском заповеднике с многолетней цикличностью осадков (табл. 3), зафиксированной там же, можно заметить полное совпадение 3- и 4-летних циклов, а также близость в средних частотах (полоса 10–15 лет). Так что численность полевки имеет здесь для подстройки близкие колебания осадков и любой из них может придавать цикличности популяции устойчивость.
Таблица 3. Соотношение величины и мощности периодических составляющих многолетней динамики плотности популяции красной полевки, некоторых климатических характеристик и цикличности плодоношения хвойных видов деревьев в Печоро-Илычском заповеднике
| Период, лет / Характеристики | 18–30 | 10–15 | 6–9 | 5–5.9 | 4–4.5 | 3.5–3.9 | 3–3.4 | 2.5–2.7 | 2–2.4 | |
| Плотность полевок | 28.4 | 10.8 | 6.0 | – | 4.4 | – | 3.1 | – | 2.4 | |
| 0.46 | 0.64 | 0.54 | 0.54 | 0.35 | 0.28 | |||||
| Количество осадков | – | 14.6 | 5.8 | 4.4 | – | 3.0 | – | 2.1 | ||
| 9.09 | 7.15 | 9.76 | 12.55 | 5.38 | ||||||
| Температура воздуха | – | 14.8 | 7.2 | 5.1 | – | – | 3.4 | 2.7 | 2.2 | |
| 0.11 | 0.16 | 0.15 | 0.11 | 0.13 | 0.10 | |||||
| Семеношение | Ель | – | 12.1 | 8.0 | – | – | 3.7 | – | 2.7 | 2.3 |
| 0.12 | 0.13 | 0.33 | 0.27 | 0.18 | ||||||
| Пихта | 24.9 | – | – | – | – | 3.5 | – | 2.7 | 2.2 | |
| 0.15 | 0.21 | 0.43 | 0.30 | |||||||
| Сосна | – | 10.9 | 5.7 | 4.1 | 3.5 | – | 2.7 | 2.1 | ||
| 0.18 | 0.16 | 0.15 | 0.14 | 0.09 | 0.24 | |||||
| Кедр | 19.3 | – | 8.3 | 5.4 | – | 3.8 | 3.1 | 2.7 | 2.2 | |
| 0.12 | 0.17 | 0.18 | 0.23 | 0.34 | 0.13 | 0.24 | ||||
Примечание. Верхняя цифра – период, лет; нижняя – мощность (амплитуда).
Фиксируемые на этой же территории многолетние изменения температуры также имели колебания, соответствующие циклам динамики красной полевки. В цикличности температуры проявлены 2- и 3-летние ритмы, которые могут поддерживать соответствующие популяционные у красной полевки. Кроме того, как и у осадков, имеется примерно 15-летний цикл, могущий затягивать 10- и 8-летний у полевки. То же может происходить и с циклами 6–7-летней в полосе частот. Так что подстройка к цикличности осадков может происходить у полевки и на средних частотах.
Обсуждение
Красная полевка среди лесных полевок наиболее семеноядна. Возможно, этим объясняется соответствие ее цикличности с цикличностью урожайности семян местных хвойных пород. Особенно точно совпадают характеристики ритмов динамики этого вида с ритмами плодоношения сосны в средних и высоких частотах. Так, характерный 10-летний цикл полевки соответствует почти такому же в динамике плодоношения сосны. Менее всего отмечена синхронность с плодоношением пихты, хотя с ее урожаем связан самый низкочастотный ритм популяции полевок (см. табл. 3). Высокочастотные популяционные ритмы вполне близки по периодам с урожайностью практически всех хвойных растений. Возможно, эти виды имеют меньшее значение как ресурс для красной полевки, чем сосна. У популяций Аляски на цикличность влияет только динамика запасов корма. Причем обилие урожая ягод там обусловлено погодными условиями (комбинации месячной летней температуры и осадков) двух предыдущих лет (Krebs et al., 2009). Так что и здесь внешним синхронизатором ритмов предполагается, хотя и опосредованно, ход климатических факторов.
Итак, местное природно-климатическое окружение предоставляет широкий диапазон колебаний, близких по периодам к циклам многолетней динамики численности красной полевки. Все они могут служить внешними ритмоводителями, придающими устойчивость популяционным циклам этого грызуна.
Сложность адаптации красной полевки заключается еще и в том, что колебания ее численности подстраиваются к одним и тем же жизненно важным внешним синхронизаторам с другими, обитающими здесь же лесными полевками (из-за значительной эколого-физиологической близости видов). Так что эти полевки, по-видимому, вынуждены разделяться во времени, чтобы снизить напряженность конкурентных отношений. Здесь тоже может пригодиться биоритмологическая основа многолетней динамики численности, ведь разделение популяций достигается еще и за счет варьирования значений фаз, периодов и мощностей циклов динамики численности (Ердаков, Панов, 1987).
Заключение
Для популяций красной полевки в каждом исследуемом регионе рассчитан весь диапазон периодических составляющих многолетней динамики численности, определены мощности циклов динамики. Гармонические составляющие спектров, построенных для разных регионов ареала красной полевки, совпадают в большинстве частотных полос. Различия в разных географических популяциях проявлены обычно только в соотношении мощностей одинаковых гармоник.
В различных регионах проявлены особенности спектров цикличности численности. Предполагается, что у северных популяций имеются низкочастотные циклы в 40, а возможно, и 60 лет. Тем не менее инвариантная часть спектра для красной полевки всегда содержит циклы в 7–10, 4.5–6, 3.5–4 и 2–3-летних полосах частот.
Популяция красной полевки подстраивает свои ритмы к соответствующим по периоду внешним циклам. Из климатических изменений это, прежде всего, цикличность осадков и, в меньшей степени, температурные циклы. Подстройка многолетних популяционных ритмов у этого вида грызунов происходит и к плодоношению хвойных в районах ее обитания. Наиболее соответствует спектр ритмов красной полевки в лишайниковых сосняках Печоро-Илычского заповедника спектру многолетней цикличности плодоношения сосны и ели.
Библиография
Бобрецов А. В., Бешкарев А. Б., Басов В. А., Васильев А. Г., Ефимов В. М., Кудрявцева Э. Н., Мегалинская И. З., Нейфельд Н. Д., Сокольский С. М., Теплов В. В., Теплов В. П. Закономерности полувековой динамики биоты девственной тайги Северного Предуралья . Сыктывкар: Госкомстат Республики Коми, 2000. 206 с.
Европейская рыжая полевка / Под ред. И. В. Башенина. М.: Наука, 1981. 351 с.
Ердаков Л. Н. Биологические ритмы: особь, популяция, сообщество. Цикличность в живых системах . LAP Lambert Academic Publishing, 2011. 152 с.
Ердаков Л. Н., Панов В. В. Циклические изменения численности лесных полевок в Барабе // Экология. 1987. № 3. С. 36–39.
Ердаков Л. Н., Савичев В. В., Чернышова О. Н. Количественная оценка популяционной цикличности у животных // Журнал общей биологии. 1990. Т. 5. № 51. С. 661–668.
Жигальский О. А., Кшнясев И. А. Популяционные циклы европейской рыжей полевки в оптимуме ареала // Экология. 2000. № 5. С. 376–383.
Ивантер Э. В., Моисеева Е. А. К экологии красной полевки (Clethrionomys rutilus Pall.) на юго-западной периферии ареала // Труды Карельского научного центра РАН. 2015. № 1. С. 37–47.
Истомин А. В. Динамика популяций и сообществ мелких млекопитающих как показатель состояния лесных экосистем (на примере Каспийско-Балтийского водораздела) . М., 2009. 481 с.
Максимов А. А., Ердаков Л. Н. Циклические процессы в сообществах животных (биоритмы, сукцессии) . Новосибирск: Наука, 1985. 236 с.
Марпл С. Л. (мл). Цифровой спектральный анализ и его приложения . М.: Мир, 1990. 584 с.
Чернявский Ф. Б., Лазуткин А. Н. Циклы леммингов и полевок на Севере . Магадан: ИБПС ДВО РАН, 2004. 150 с.
Bjørnstad O. N., Champely S., Stenseth N. C., Saitoh T. Cyclicity and stability of grey-sided voles, Clethrionomys rufocanus, of Hokkaido: spectral and principal components analyses // Philos. Trans. Biol. Sci. 1996. Vol. 351. P. 867–875.
Boonstra R., Krebs C. J. Population dynamics of red-backed voles (Myodes) in North America // Oecologia. 2012. Vol. 168. P. 601–620.
Fuller W. A. Demography of a subarctic population of Clethrionomys gapperi : numbers and survival // Can. J. Zool. 1977. Vol. 55. № 1. P. 42–51.
Hanski I., Turchin P., Korpimaki E., Henttonen H. Population oscillations of boreal rodents: regulation by mustelid predators leads to chaos // Nature. 1993. Vol. 364. № 6434. P. 232–235.
Hansson L., Henttonen H. Regional differences in cyclicity and reproduction in Clethrionomys species: Are they related? // Ann. Zool. Fennici. 1985. Vol. 22. P. 277–288.
Hansson L., Stenseth N. C. Modelling small rodent population dynamics: suggestions to empiricists, theoreticians and editors // Oikos. 1988. Vol. 52. № 2. P. 227–229.
Henttonen H., McGuire A. D., Hansson L. Comparisons of amplitudes and frequencies (spectral analyses) of density variations in long-term data sets of Clethrionomys species // Ann. Zool. Fennici. 1985. Vol. 22. P. 221–227.
Krebs C. J., Boonstra R., Cowcill K., Kenneya A. J. Climatic determinants of berry crops in the boreal forest of the southwestern Yukon // Botany. 2009. Vol. 87. № 4. P. 401–408.
McArdle B. H., Gaston K. J. Comparing population variabilities // Oikos. 1992. Vol. 64. № 3. P. 610–612.
Saitoh T., Bjørnstad O. N., Stenseth N. C. Density dependence in voles and mice: a comparative study // Ecology. 1999. Vol. 80. № 2. P. 638–650.
Stenseth N. C. Mathematical models of Microtine cycles: Models and the real world // Acta Zool. Fenn. 1985. Vol. 173. P. 7–12.
Stenseth N. C., Bjørnstad O. N., Saitoh T. A gradient from stable to cyclic populations of Clethrionomys rufocanus in Hokkaido, Japan. // Proceedings. Biol. Sci. 1996. Vol. 263. № 1374. P. 1117–1126.
Turchin P., Hanski I. An empirically based model for latitudinal gradient in vole population dynamics // Am. Nat. 1997. Vol. 149. № 5. P. 842–874.
West S. D. Dynamics of colonization and abundance in Central Alaskan populations of the northern red-backed vole, Clethrionomys rutilus // J. Mammal. 1982. Vol. 63. № 1. P. 128–143.

 © 2011 - 2024
© 2011 - 2024