



Введение
Гнездовой паразитизм у птиц, и облигатный, и факультативный, хорошо известное исследователям явление. Его рассматривают как альтернативную репродуктивную тактику, при которой паразитирующая самка откладывает яйца в чужие гнезда, не насиживает их и заботу о потомстве оставляет хозяину гнезда. Гнездовой паразитизм особенно широко распространен среди гусеобразных птиц (Sorenson, 1992; Geffen, Yom-Tov, 2001) и за исключением одного южноамериканского вида, черноголовой древесной утки (Heteronetta atricapilla), проявляется в факультативной форме и может быть как внутри-, так и межвидовым. Случаи гнездового паразитизма зарегистрированы у более половины видов этого отряда, обитающих в различных регионах, гнездящихся в открытых и в лесных местообитаниях, отдельными парами и колониально. В одни списки включены 76 видов (Geffen, Yom-Tov, 2001), в другие – 88 (Нумеров, 2003), и по мере поступления новых данных о малоизученных видах, скорее всего, число видов увеличится. Опубликованы всесторонние обзоры особенностей гнездового паразитизма у разных видов водоплавающих птиц и гипотез его происхождения (Sayler, 1992; Нумеров, 2003; Pöysä et al., 2014). Основной акцент делается на исследовании либо только внутривидового гнездового паразитизма, и таких работ большинство, либо только межвидового. Одновременное изучение внутривидового и факультативного межвидового гнездового паразитизма позволяет оценить значение этих форм поведения в жизни популяций.
Цель нашей работы – дать количественную оценку внутривидового и межвидового гнездового паразитизма у уток, гнездящихся на оз. Кротово, а также их межгодовой динамики в зависимости от относительной численности каждого вида в утином сообществе.
Материалы
Материал по гнездовому паразитизму собирался в ходе исследований экологии утиных птиц в сезоны размножения с 1970 по 2016 г. на оз. Кротово (53°72' с. ш., 77°88' в. д.) в Карасукском районе Новосибирской области. Озеро Кротово, площадь которого (345–485 га) изменяется в зависимости от обводненности, входит в Карасукский озерно-займищный район, занимающий низовья бассейна р. Карасук. Оно относится к озерам с внутриозерно-сплавинным типом зарастания (Волгин, Сипко, 1982). Сплавина и заросли надводной растительности занимают около 60 % площади озера, благодаря чему на озере имеются хорошие условия для гнездования уток. Основа растительности сплавин – тростник (Phragmites communis) и папоротник (Dryopteris thelypteris). Подробно место работы описано ранее (Михантьев, Селиванова, 2008).
За исключением 6 лет (1976, 1977, 1980, 2012, 2013, 2014) утиные гнезда учитывали с начала мая до конца июля методом сплошного осмотра островов и кромки сплавины на постоянных площадках, расположенных по периметру озера, общей площадью примерно 28 га. Периодичность повторных учетов и проверок гнезд 7–10 дней. Яйца измеряли, описывали цвет скорлупы, определяли стадию инкубации по водяному тесту (Westerskov, 1950; Меднис, Блум, 1976) и метили водостойким маркером.
На озере найдены гнезда 11 видов гусеобразных (Михантьев, Селиванова, 2010), основные из них – три вида речных уток: кряква (Anas platyrhynchos), серая утка (A. strepera), широконоска (A. clypeata), и три вида нырковых уток: красноголовый нырок (Aythya ferina), хохлатая чернеть (A. fuligula), красноносый нырок (Netta rufina). Было обследовано 6654 гнезда (табл. 1). В гнездах каждого вида уток находили чужие яйца, и каждый вид в той или иной степени выступал в роли паразита (табл. 2).
Таблица 1. Число утиных гнезд и смешанных кладок, учтенных на оз. Кротово (1970–2016 гг.)
Table 1. Number of duck nests and mexed cluthces counted on Lake Krotovo and number of nests with parasitic eggs (1970–2016)
| Виды Species | Число гнезд No. nests | Найдены не разоренными found not depredated | Содержали чужие яйца with parasitic eggs | % |
| Ay. ferina | 1815 | 1487 | 483 | 32,5 |
| Ay. fuligula | 2153 | 1774 | 515 | 29.0 |
| N. rufina | 49 | 45 | 12 | 26.7 |
| A. platyrhynchos | 2227 | 1660 | 249 | 15.0 |
| A. strepera | 323 | 280 | 88 | 31.4 |
| A. clypeata | 40 | 40 | 13 | 32.5 |
| Другие | 47 | 47 | 15 | 31.9 |
| Всего | 6654 | 5333 | 1375 | 25.8 |
Таблица 2. Число яиц, отложенных утками в чужие гнезда (оз. Кротово, 1970–2016 гг.)
Table 2. The number of parasitic eggs laid by ducks (in rows) into host species’ nests (in columns) on Lake Krotovo, 1970–2016
| Вид | Всего яиц N | Из них паразитических яиц, отложенных в гнезда вида-хозяина: | % | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | другие | сумма | |||
| A. ferina (1) | 12863 | 1655 | 235 | 17 | 258 | 61 | 9 | 27 | 2262 | 17.6 |
| A. fuligula (2) | 14516 | 172 | 1714 | 6 | 124 | 128 | 15 | 43 | 2202 | 15.2 |
| N. rufina (3) | 481 | 53 | 22 | 8 | 43 | 4 | 0 | 3 | 133 | 27.7 |
| A. platyrhynch.(4) | 12095 | 27 | 12 | 1 | 134 | 15 | 2 | 0 | 191 | 1.6 |
| A. strepera (5) | 2259 | 12 | 121 | 0 | 31 | 21 | 0 | 1 | 186 | 8.2 |
| A. clypeata (6) | 307 | 0 | 8 | 0 | 9 | 0 | 1 | 0 | 18 | 5.9 |
| Всего | 42521 | 1919 | 2112 | 32 | 599 | 229 | 27 | 74 | 4992 | 11.7 |
Примечание. Жирным шрифтом выделено число паразитических яиц в конспецифичных кладках.
Number of eggs laid intraspecifically mark in bold.
Методы
Межвидовые смешанные кладки, как правило, хорошо определяются по различиям размеров, формы и окраски яиц. Внутривидовые смешанные кладки определяли как по отличию размеров, формы и цвета яиц, так и по другим широко применяемым критериям (Меднис, 1968; Dugger, Blums, 2001; Нумеров, 2003). А именно: по появлению более одного яйца в день на стадии откладки яиц; по появлению новых яиц спустя два и более дня после начала насиживания; по нахождению в гнезде яиц на сильно различающихся стадиях инкубации; по величине кладки, превышающей 14 яиц; по оставшимся в гнезде после ухода выводка яйцам с живыми недоразвитыми эмбрионами. На смешанную кладку указывали и яйца, разбросанные около гнезда. Вероятно, они были отложены около гнезда паразитирующей особью в случае нахождения на гнезде самки-хозяйки либо выкатились в ходе конфликта между самками.
Соотношение видов определяли по всем учтенным гнездам (см. табл. 1). При расчетах показателей гнездового паразитизма не учитывали гнезда, разоренные до их обнаружения (1321 гн.), поскольку было невозможно точно определить наличие в них чужих яиц.
Прежде чем перейти к изложению результатов, необходимо дать определения применяемых терминов. Экстенсивность паразитизма («степень паразитирования») – отношение числа гнезд, содержащих чужие яйца, к общему числу обследованных гнезд этого вида в процентах. Интенсивность паразитизма – число яиц или птенцов вида-паразита в одном гнезде хозяина (Нумеров, 2003, стр. 9). Склонность вида к гнездовому паразитизму можно было бы оценить относительным числом самок, отложивших яйца в чужие гнезда, от общей численности самок этого вида, при условии установления принадлежности этих яиц конкретной самке. Однако это невозможно сделать без применения генетических и биохимических методов. Мы предлагаем оценивать склонность вида к паразитизму (или уровень гнездового паразитизма) процентным отношением числа яиц, отложенных в чужие гнезда своего и других видов, к общему числу яиц этого вида. Выбирает ли паразитирующий вид гнезда какого-то определенного вида уток, оценивали коэффициентом предпочтения. Коэффициенты рассчитывали для каждого года путем деления долей паразитических яиц в кладках своего и других видов соответственно на доли гнезд этих видов в утином сообществе на озере.
Весь многолетний материал занесен в базу данных в Microsoft Access, там создавались таблицы для расчетов. Статистическая оценка результатов была проведена с использованием пакета программ PAST: проверка на нормальное распределение, параметрические и непараметрические анализы. Также для статистических анализов и построения графиков использовали Microsoft Excel. Все средние величины приведены с ошибкой (±SE). Для проверки различий средних значений применяли двухвыборочный t-критерий Стьюдента с различными дисперсиями и U-критерий Манна – Уитни. Для измерения степени сопряженности уровня паразитизма с численностью пользовались коэффициентом корреляции Пирсона (r) и коэффициентом ранговой корреляции Спирмена (rs).
Результаты
На гнездовании на оз. Кротово преобладали по численности кряква, красноголовый нырок и хохлатая чернеть. Обычна на гнездовании серая утка. Остальные виды гнездились единичными парами и не ежегодно. Соотношение видов сильно варьировало по годам.
За все время исследования зафиксирован 851 случай внутривидового и 666 случаев межвидового гнездового паразитизма, всего 1517. С учетом случаев паразитирования в одном гнезде нескольких видов уток, гнезд, содержащих яйца не только хозяйки гнезда, было всего 1375. Из них: 1240 гнезд (23.2 %) содержали чужие яйца какого-то одного вида, 128 (2.4 %) – двух видов и 7 гнезд (0.1 %) – трех видов уток.
Рассмотрим участие разных видов в процессе гнездового паразитизма как в качестве жертв гнездового паразитизма, так и в качестве паразитов.
Кряква. На оз. Кротово это доминирующий вид уток. Число ее гнезд, найденных в разные годы, колебалось от 10 до 129, в среднем было 54.3 ± 4.8 гнезда. В связи с колебаниями численности всех видов уток изменялась и доля кряквы в утином сообществе: от 10.6 до 77.5 %, среднегодовая доля равна 37.1 ± 2.5 %.
Суммарная экстенсивность паразитизма в гнездах кряквы составила 15.0 %. Чаще других в ее гнезда откладывали яйца красноголовые нырки (8.1 %) и хохлатые чернети (4.1 %). Около 3 % кладок содержали яйца малочисленных видов: красноносого нырка, серой утки и широконоски. Внутривидовой паразитизм у кряквы выявлен в 2.4 % гнезд. Иногда в одном гнезде находили чужие яйца двух и трех видов уток. В 11 случаях это были яйца красноголового нырка и хохлатой чернети, в 3 случаях – красноголового и красноносого нырка, в 4 случаях – хохлатой чернети и кряквы. Дважды в гнездах кряквы отмечено совместное паразитирование хохлатой чернети и серой утки, хохлатой чернети и широконоски. По одному гнезду кряквы содержали чужие яйца трех видов уток: красноголового нырка, хохлатой чернети и широконоски; красноголового нырка, хохлатой чернети и серой утки. В среднем в одной смешанной кладке кряквы было 2.4 яйца, не принадлежащих хозяйке гнезда.
В качестве гнездового паразита крякву регистрировали редко. Ее яйца нашли всего в 75 гнездах (1.4 % от общего числа утиных гнезд), из которых в 40 гнездах (53.3 %) установлен внутривидовой паразитизм. Яйца кряквы отмечены в 16 гнездах красноголового нырка, в 11 – хохлатой чернети, в 6 – серой утки (21.3, 14.7 и 8 % соответственно). А также в одном гнезде красноносого нырка и в одном гнезде широконоски (по 1.3 %).
По сравнению с другими видами уток доля яиц кряквы, отложенных в чужие гнезда, мала (см. табл. 2). Она изменялась по годам от 0 до 6.7 %, составив в среднем 1.5 ± 0.3 %. Интенсивность гнездового паразитизма кряквы во внутривидовых и межвидовых смешанных кладках была от 1 до 10 яиц. Большая часть этих кладок (81.3 %) содержали не более 4 чужих яиц кряквы.
Красноголовый нырок – на оз. Кротово многочисленный вид. В среднем в год учитывали 44.3 ± 4.7 (от 2 до 140) гнезда. Доля гнезд красноголового нырка от общего числа утиных гнезд изменялась по годам от 4.5 до 57.5 % (26.6 ± 1.9 %).
Суммарная экстенсивность паразитизма в гнездах красноголового нырка была высокой (32.5 %) за счет высокой экстенсивности внутривидового паразитизма (26.6 %). В ряде случаев установлено паразитирование красноголового нырка вместе с хохлатой чернетью (15 гнезд), красноносым нырком (9 гн.), кряквой (3 гн.), серой уткой (1 гн.), хохлатой чернетью и красноносым нырком (1 гн.). Яйца хохлатой чернети были найдены в 4.8 % гнезд красноголового нырка. Кроме случаев совместного паразитирования хохлатой чернети с красноголовым нырком, зафиксированы 3 случая с красноносым нырком и 1 с кряквой. В 27 (2.8 %) гнездах красноголового нырка были яйца красноносого нырка, в 16 (1.1 %) – кряквы и в 6 (0.4 %) – серой утки. В среднем в одной смешанной кладке красноголового нырка было 4.0 чужих яйца.
В качестве гнездового паразита красноголовый нырок определен в 691 гнезде (13.0 % от общего числа утиных гнезд), больше половины (57.3 %) из которых были гнезда своего вида. Остальные – других видов уток, а именно: кряквы (19.4 %), хохлатой чернети (15.3 %), серой утки (4.6 %), красноносого нырка (1.0 %), широконоски (0.9 %) и др. (1.4 %).
В абсолютном выражении сумма яиц красноголового нырка, отложенных в чужие гнезда, больше, чем у остальных видов (табл. 2). Его яйца ежегодно регистрировали в чужих гнездах. Доля таких яиц изменялась по годам от 2.7 до 39.4 % и в среднем составляла 17.2 ± 1.3 %. Интенсивность гнездового паразитизма была от 1 до 20 яиц, но в большинстве случаев (76.0 %) от 1 до 4 яиц.
Хохлатая чернеть. Как и у предыдущих видов, у хохлатой чернети происходили колебания численности популяции, что отразилось на числе ежегодно найденных гнезд (от 2 до 130). Средний показатель равен 51.3 ± 5.6 гнезда. Доля гнезд этих нырков в сообществе утиных колебалась от 9.1 до 47.4 % (29.4 ± 1.7 %) в результате изменения численности всех видов уток.
Суммарная экстенсивность паразитизма в ее гнездах составила 29.0 %, из которых 22.7 % содержали конспецифичные яйца. В их числе 37 случаев паразитирования хохлатой чернети вместе с красноголовым нырком, 4 – с кряквой, 2 – с серой уткой, 1 – с широконоской и 1 – с красноносым и красноголовым нырками. Яйца красноголового нырка отмечены в 106 (6.0 %) гнездах хохлатой чернети. Из них в 65 гнездах были чужие яйца только этого вида, а в остальных, кроме упомянутого паразитирования с хохлатой чернетью, было по одному случаю с красноносым нырком, с серой уткой и с широконоской. В 34 (1.9 %) гнездах были яйца серой утки. Экстенсивность паразитизма остальными видами составила менее процента. Одно гнездо хохлатой чернети со смешанной кладкой в среднем содержало 4.1 чужих яйца.
Хохлатая чернеть в свою очередь сама часто откладывала яйца в чужие гнезда. Ее яйца обнаружены в 616 гнездах разных видов уток (11.6 % от общего числа гнезд). Конспецифичные смешанные кладки составили 65.4 %. Примерно с одинаковой частотой хохлатая чернеть откладывала яйца в гнезда красноголового нырка и кряквы — 11.7 и 11.0 % соответственно. Относительно часто хохлатая чернеть откладывала яйца в гнезда серой утки, численность которой в 5–6 раз ниже, чем у хохлатой чернети, красноголового нырка и кряквы. Ее яйца обнаружены в 49 (8.0 %) гнездах серой утки. Еще 24 случая паразитирования хохлатой чернети зафиксированы в гнездах красноносого нырка (0.5 %), широконоски (1.3 %) и других малочисленных видов уток (2 %).
Доля яиц, отложенных в чужие гнезда, изменялась по годам от 0 до 38.2 % и в среднем была 13.4 ± 1.3 %. Интенсивность гнездового паразитизма хохлатой чернети колеблется от 1 до 22 яиц в одном гнезде, чаще всего (73.9 %) не более 4 яиц.
Серая утка гнездится на оз. Кротово постоянно, но численность ее не бывает высокой. Ежегодно наблюдали от 1 до 26 (в среднем 7.9 ± 1.1) гнезд. Доля серой утки в утином сообществе в разные годы изменялась от 0.9 до 10.4 % гнезд. Среднегодовой показатель – 4.8 ± 0.4 %.
Суммарная экстенсивность паразитизма в гнездах серой утки была высокой (31.4 %). В 17.5 % гнезд были яйца хохлатой чернети, в 11.4 % – красноголового нырка. В небольшом числе гнезд обнаружены яйца кряквы (2.1 %) и красноносого нырка (1.0 %). Внутривидовой гнездовой паразитизм отмечен в 3.2 % случаев. Зафиксировано 10 фактов совместного паразитирования двумя и более видами уток. Пять гнезд содержали яйца красноголового нырка и хохлатой чернети, по одному гнезду – хохлатой чернети и кряквы; хохлатой чернети и серой утки; хохлатой чернети и шилохвости; красноголового нырка и серой утки. В одном гнезде нашли чужие яйца трех видов: красноголового нырка, хохлатой чернети и серой утки. В одной смешанной кладке серой утки в среднем было 2.6 чужих яица.
Паразитизм серой уткой зарегистрирован 65 раз (1.2 % от числа всех утиных гнезд на озере). Чаще всего она подкладывала яйца в гнезда хохлатой чернети – 52.3 % от всех случаев ее паразитизма, реже в гнезда кряквы (24.6 %). Единичные встречи яиц серой утки были зарегистрированы в конспецифичных гнездах (13.8 %) и в гнездах красноголового нырка (9.2 %).
В условиях озера Кротово, в отличие от кряквы, красноголового нырка и хохлатой чернети, серая утка подкладывала меньше яиц в конспецифичные, чем в интерспецифичные кладки. Несмотря на низкую численность этой утки, ее яиц в чужих гнездах зарегистрировано столько же, сколько и у кряквы (см. табл. 2). Доля яиц, отложенных в чужие гнезда, колебалась по годам от 0 до 57.1 %, составив в среднем 9.4 ± 2.1 %. Интенсивность паразитизма была от 1 до 12 яиц, но 81.8 % гнезд содержали не более 4 ее яиц.
Красноносый нырок гнездится на озере не ежегодно и в небольшом числе (максимум 6 гнезд). Максимальное значение доли его гнезд в утином сообществе 4.5 %, а среднегодовое – 0.7 ± 0.2 %.
В 26.7 % кладок красноносого нырка были чужие яйца либо конспецифичных особей (4.4 %), либо уток других видов. Яйца красноголового нырка были обнаружены в 6 гнездах (13.3 %), яйца хохлатой чернети – в двух (4.4 %), вместе яйца этих двух видов – в одном гнезде (2.2 %). В одном гнезде были яйца кряквы. В среднем в одной смешанной кладке красноносого нырка было 2.7 чужих яйца.
Паразитизм красноносого нырка в чужих гнездах зарегистрирован 56 раз. В большинстве случаев (48.2 %) это были гнезда красноголового нырка. Также яйца красноносого нырка были в 16 гнездах кряквы (28.6 %), в 7 гнездах хохлатой чернети (12.5 %), в двух гнездах серой утки (3.6 %) и в двух гнездах других редко гнездящихся на озере видов (3.6 %). Конспецифичный гнездовой паразитизм красноносого нырка был отмечен дважды (3.6 %).
Более четверти от общего количества яиц красноносого нырка отложены в чужие гнезда (табл. 2). Особо следует отметить, что даже в годы, когда гнезд этого нырка не находили, яйца, принадлежащие ему, регистрировали в гнездах разных видов уток. Доля яиц, отложенных в чужие гнезда, изменялась по годам от 0 до 100 % (в среднем 29.6 ± 6.4 %). Интенсивность паразитизма была от 1 до 10 яиц, 90.7 % гнезд содержали не более 4 яиц этого вида уток.
Широконоска, как и красноносый нырок, малочисленный и не ежегодно гнездящийся на озере вид. Находили ее в ходе 24 сезонов размножения и максимум 5 гнезд. Доля гнезд широконоски в утином сообществе достигала 8.3 %, среднегодовая доля равна 0.7 ± 0.2 %.
Всего мы нашли 40 гнезд широконоски, из них 13 (32.5 %) содержали чужие яйца: хохлатой чернети (12.5 %), красноголового нырка (10 %) и вместе этих двух видов (5 %). В одно (2.5 %) гнездо подложили яйца хохлатая чернеть и кряква и в одно – широконоска. В смешанной кладке широконоски было в среднем 2.1 чужих яйца.
Яйца широконоски были обнаружены всего в 9 чужих гнездах: в четырех гнездах хохлатой чернети, в четырех кряквы и в одном своего вида. Доля яиц, отложенных в чужие гнезда, колебалась по годам от 0 до 28.6 %, составив в среднем 7.1 ± 3.1 %. Интенсивность паразитизма была от 1 до 5 яиц, 88.9 % гнезд содержали от 1 до 4 паразитических яиц этого вида.
Таким образом, собранные нами фактические данные свидетельствуют о разном уровне гнездового паразитизма у уток. Нырковые утки откладывают яиц в чужие гнезда в среднем на порядок больше, чем речные. Полученное распределение по уровню гнездового паразитизма (т. е. по ежегодной доле яиц, отложенных в чужие гнезда) соответствует нормальному у красноголового нырка (χ² = 1.2, р = 0.27), хохлатой чернети (χ² = 0.4, р = 0.53), красноносого нырка (χ² = 2.3, р = 0.12) и кряквы (χ² = 2.9, р = 0.1), но не подтверждается для серой утки и широконоски (р < 0.05). Поскольку соответствие нормальному распределению – это необходимое требование для применения параметрического критерия Стьюдента, мы дополнительно сравнили выборки непараметрическим критерием. Средние уровни паразитизма красноголового нырка (17.2 ± 1.3 %) и красноносого нырка (29.6 ± 6.4 %) значимо не различались и были достоверно выше (t > 2.05, р < 0.04), чем у остальных видов. Хохлатая чернеть (13.4 ± 1.3 %) и серая утка (9.4 ± 2.1 %) по этому показателю достоверно (t > 3.57, р < 0.001) опережали крякву (1.5 ± 0.3 %). Среднегодовая доля паразитических яиц широконоски (7.1 ± 3.1 %) достоверно (t > 2.85, р < 0.008) отличалась только от нырковых уток. Сравнение выборок U-критерием Манна – Уитни подтвердило достоверность этих различий, а также выявило значимое различие между серой уткой и хохлатой чернетью (z = –3.3, р < 0.0008), серой уткой и широконоской (z = –2.3, р < 0.019).
Преобладающие по численности на озере кряквы, красноголовые нырки и хохлатые чернети достоверно чаще подкладывали яйца в гнезда своего вида (табл. 3). Немногочисленные виды (красноносый нырок, серая утка) чаще подкладывали яйца другим видам уток, что можно объяснить недостатком конспецифичных гнезд. Различия уровней конспецифичного и интерспецифичного паразитизма у широконоски не выявлено. Распределения данных отдельно по уровням внутривидового и межвидового гнездового паразитизма соответствуют нормальному только у красноголового нырка и хохлатой чернети (χ² < 3.4, р > 0.07) и не подтверждаются для остальных видов.
Таблица 3. Относительное число паразитических яиц отдельных видов уток в конспецифичных (1) и интерспецифичных (2) кладках от общего числа яиц, отложенных видом, оз. Кротово, 1970–2016 гг., M ± SE
Table 3. Proportions of all (%) eggs laid parasitically in the nests of conspecifics (1) and other duck species (2), Lake Krotovo, 1970–2016, M ± SE
| Паразитирующий вид Parasitic Species | 1 | 2 | t-критерий
t-test |
U-критерий U-test | ||
| % | % | t | p | z | p | |
| A. ferina | 11.2 ± 1.0 | 6.0 ± 0.9 | 3.8 | 0.0003 | –4.1 | <10-4 |
| A. fuligula | 10.1 ± 1.0 | 3.3 ± 0.5 | 6.0 | <10-5 | –4.5 | <10-5 |
| N. rufina | 1.5 ± 1.1 | 28.1 ± 6.5 | 4.0 | 0.0005 | –4.2 | <10-4 |
| A. platyrhynchos | 1.1 ± 0.3 | 0.4 ± 0.1 | 2.5 | 0.016 | –1.9 | 0.048 |
| A. strepera | 0.6 ± 0.2 | 9.0 ± 2.1 | 3.8 | 0.0004 | –4.7 | <10-5 |
| A. clypeata | 0.24 ± 0.19 | 6.9 ± 3.1 | 2.04 | 0.053 | –1.8 | 0.07 |
Корреляционный анализ между уровнем гнездового паразитизма и числом гнезд каждого вида на озере по годам выявил несколько достоверных зависимостей у массовых видов уток (табл. 4). У красноголового нырка доля паразитических яиц во внутривидовых кладках увеличивалась, в межвидовых уменьшалась, а суммарная доля не зависела от численности. У хохлатой чернети общая доля паразитических яиц достоверно возрастала с увеличением числа ее гнезд на озере, причем главным образом за счет внутривидового паразитизма. У кряквы прослеживалась тенденция повышения уровня и внутривидового, и межвидового паразитизма с увеличением числа гнезд. Уровень гнездового паразитизма серой утки не зависел от численности.
Таблица 4. Оценка сопряженности доли паразитических яиц отдельных видов уток в конспецифичных (1), интерспецифичных (2) смешанных кладках и их суммы (3) с числом их гнезд на оз. Кротово, 1970–2016 гг.
Table 4. Correlation coefficients for the proportion of parasitic eggs (considered in comparative relation to all eggs) laid intraspecifically (1), laid interspecifically (2), and total (3) versus nest numbers of this species on Lake Krotovo, 1970–2016
| r / rs | ||||
| Паразитирующий вид Parasitic species | 1 | 2 | 3 | n |
| A. ferina | 0.45**/0.51*** | -0.32*/-0.30 | 0.18/0.29 | 41 |
| A. fuligula | 0.52***/0.56*** | 0.25/0.31 | 0.60***/0.65*** | 41 |
| A. platyrhynchos | 0.21/0.46** | 0.30/0.59*** | 0.28/0.49** | 41 |
| A. strepera | 0.20/0.31 | -0.18/0.15 | -0.12/0.26 | 41 |
Примечание. * – P < 0.05; ** – P < 0.01; *** – P < 0.001
Для дальнейшего анализа у каждого вида выбраны только те годы, когда зафиксированы случаи паразитизма. Процент паразитических яиц в конспецифичных кладках рассчитывали по годам от общей суммы яиц в чужих гнездах. Проанализировали изменение доли таких яиц у четырех массовых видов уток при изменении доли гнезд каждого вида относительно общего числа гнезд на озере. Установили, что при увеличении доли гнезд красноголового нырка достоверно возрастала и доля его паразитических яиц во внутривидовых смешанных кладках (рисунок; r = 0.61, rs = 0.66, p < 0.001, n = 41). Немного слабее корреляция у серой утки (r = 0.48, rs = 0.43, p < 0.05, n = 27). У хохлатой чернети эта тенденция недостоверна (r = 0.27, rs = 0.20, p < 0.1, n = 39), а у кряквы – отсутствует (рис.).
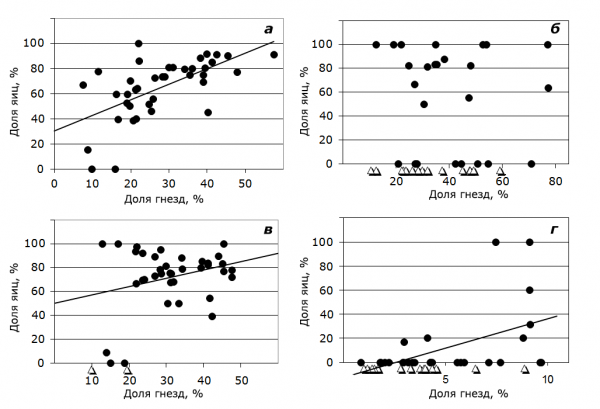
Рис. Соотношение доли паразитических яиц, отложенных в гнезда своего вида, и доли гнезд этого вида в утином сообществе у а – красноголового нырка, б – кряквы, в – хохлатой чернети, г – серой утки (оз Кротово, 1970–2016 гг.): ∆ – отмечены значения, когда вид не паразитировал. По оси Х процент гнезд данного вида от общего числа утиных гнезд в год. По оси Y доля внутривидовых от общего числа паразитических яиц данного вида
Fig. Relationship of the proportion of intraspecific parasitic eggs with the proportion of each species in the duck community: а – pochard, б – mallard, в – tufted duck, г – gadwall (Lake Krotovo, 1970–2016): ∆ – no parasitism of this species was observed. The X axis shows the percentage of nests of a given species of the total number of duck nests by years. The Y axis shows the proportion of intraspecific parasitic eggs and the total number of those of a given species
Остается вопрос: действует ли паразитирующий вид избирательно, откладывая яйца в чужие гнезда? С целью ответа на него были рассчитаны коэффициенты предпочтения. Например, в 2016 году красноголовые нырки отложили 143 паразитических яйца: 87.4 % – своему виду, 6.3 % – крякве, по 2.8 % – хохлатой чернети и красноносому нырку и 0.7 % – остальным видам уток. Найдено 200 гнезд: 57.5 % – гнезда красноголового нырка, 21 % – кряквы, 14 % – хохлатой чернети, 2 % – красноносого нырка и 5.5 % – гнезда остальных видов. Коэффициент предпочтения конспецифичных гнезд – 1.5, гнезд кряквы – 0.3, хохлатой чернети – 0.2, красноносого нырка – 1.4 и остальных видов – 0.1.
Используя для расчета коэффициентов предпочтения массив данных за все годы, мы получили следующие результаты. Самки красноголового нырка (z < –4.7, р < 10-5), хохлатой чернети (z < –4.5, р < 10-4) и кряквы (z < –2.2, р < 0.03) предпочитали паразитировать в гнездах своего вида, между гнездами других видов они не делали различий (табл. 5). Серая утка предпочитала гнезда своего вида и хохлатой чернети (z < –2.1, р < 0.03). По причине малочисленности красноносых нырков и широконосок нельзя дать достоверного заключения, гнезда каких видов они предпочитают, но тенденция выбора гнезд своего вида прослеживается и у них (см. табл. 5).
Таблица 5. Коэффициенты предпочтения видом-паразитом гнезд вида-хозяина (M ± SE), оз. Кротово, 1970–2016 гг.
Table 5. The quotient of the share of parasitic eggs in host nest (considered in comparative relation to all parasitic eggs of parasitic species) to the proportion of nests of host species within the breeding duck community on Krotovo Lake, 1970–2016 (M ± SE)
| Вид-паразит Parasitic species | Вид-хозяин Host species | |||||
| A. ferina | A. fuligula | N. rufina | A. platyrh. | A. strepera | A. clypeata | |
| A. ferina | 2.4 ± 0.2 | 0.3 ± 0.1 | 0.6 ± 0.3 | 0.4 ±0.1 | 0.8 ± 0.2 | 0.7 ± 0.3 |
| A. fuligula | 0.4 ±0.1 | 2.4 ± 0.2 | 0.4 ± 0.2 | 0.2 ± 0.1 | 1.0 ± 0.2 | 0.3 ± 0.2 |
| N. rufina | 0.8 ± 0.3 | 0.9 ± 0.3 | 9.9 ± 6.2 | 0.7 ± 0.2 | 0.6 ± 0.4 | – |
| A. platyrhynchos | 0.6 ±0.2 | 0.4 ±0.2 | 0.6 ± 0.4 | 1.9 ± 0.3 | 0.7 ± 0.4 | 1.2 ± 0.9 |
| A. strepera | 0.3 ±0.1 | 1.6 ± 0.2 | – | 0.7 ± 0.2 | 1.7 ± 0.6 | – |
| A. clypeata | – | 1.1 ± 0.6 | – | 1.3 ± 0.6 | – | 2.3 ± 1.5 |
Примечание. Жирным шрифтом выделены коэффициенты предпочтения конспецифичных гнезд.
Intraspecific quotients mark in bold.
Обсуждение
Наши многолетние наблюдения свидетельствуют о широком распространении у уток гнездового паразитизма, как внутри-, так и межвидового, что согласуется со сведениями, имеющимися в литературе (Нумеров, 2003). Уровни паразитизма значительно различаются у речных и нырковых уток. Их оценки, полученные нашими методами, возможно, занижены, но они мало отличаются от результатов работ с применением более точных биохимических и генетических методов. Эти работы, как правило, краткосрочные, 1–3 года, выполнены на небольшом материале, 25–40 гнезд, в местах скученного гнездования уток. Так, в кладках красноголового нырка в плотных островных поселениях на искусственных рыборазводных прудах 37.5–39 % яиц содержались во внутривидовых смешанных кладках (Petrželková et al., 2013; Šťovíček et al., 2013). В гнездах кряквы, по данным Я. Крейсингера с соавторами (Kreisinger et al., 2010), выводилось до 10 % утят других самок кряквы. У серой утки в течение одного гнездового сезона не выявлено случаев интраспецифичного гнездового паразитизма (Peters et al., 2003).
Многие исследователи связывают уровень внутривидового гнездового паразитизма у водоплавающих птиц с численностью или плотностью гнездования. Большинство этих исследований посвящены видам, гнездящимся либо в дуплах, либо колониально (Нумеров, 2003; Eadie et al., 1998; Waldeck et al., 2004; Roy et al., 2009; Deng et al., 2011). Есть сообщения о существовании такой связи у красноголового нырка и хохлатой чернети (Яновский, Богдановская, 1982; Sukhanova, 1996), у кряквы и серой утки (Duebbert et al., 1983; Hines, Mitchell, 1984). В ряде работ содержатся сведения об усилении межвидового гнездового паразитизма у уток с ростом численности и плотности гнездования (Lokemoen, 1991; Musil, Neužilova, 2009). Наши данные подтверждают достоверную зависимость уровня внутривидового паразитизма от численности у красноголового нырка, хохлатой чернети и кряквы, а общего уровня паразитизма только у двух последних.
Факультативный межвидовой гнездовой паразитизм у выводковых видов птиц эволюционно поддерживается теми же механизмами, что и внутривидовой гнездовой паразитизм. В частности, обеспечивается более широкий выбор гнезд потенциальных хозяев в течение более длительного периода (Lyon, Eadie, 1991; Beauchamp, 1998). Мы установили, что в условиях оз. Кротово у малочисленных видов преобладает межвидовой паразитизм, а у массовых – внутривидовой. Эта закономерность отмечалась в других регионах. При падении численности отдельных видов уток повышался уровнь межвидового паразитизма (Lebedeva, Markitan, 2001; Musil, Neužilova, 2009). Преимущественно внутривидовой паразитизм регистрировался у массовых видов уток (Hines, Mitchell, 1984; Amat, 1993; Пыжьянов, Березовская, 2010).
Самки кряквы, менее других склонные к гнездовому паразитизму, вероятно, в соответствии с гипотезой «Лучше хоть что-то» (Best-of-a-bad-job) (Lyon, Eadie, 2008), откладывают яйца в чужие гнезда тогда, когда у них нет возможности гнездиться самостоятельно. Причиной этого могут быть ряд средовых и физиологических факторов, например нехватка территорий и мест для устройства гнезд, состояние организма, возраст и недостаток опыта, потеря гнезда, отсутствие постоянного селезня. На оз. Кротово подкладывание яиц кряквой в чужие гнезда отмечалось не ежегодно. Чаще преобладал внутривидовой паразитизм, но как при низкой доле гнезд кряквы в утином сообществе, так и при высокой были годы, когда больше паразитических яиц откладывалось в гнезда других видов. Кряква – высоко территориальный вид (Михантьев, 1980). На оз. Кротово плотность ее гнездования не достигала значений, приводящих к такому повышению уровня гнездового паразитизма (до 20.6 % внутривидовых смешанных кладок), как было показано на примере переуплотненного гнездования крякв в искусственно созданных условиях с подкормкой, электрической изгородью, защищающей от наземных хищников, и искусственными гнездами (Titman, Lowther, 1975).
Нырковые утки паразитируют в чужих гнездах чаще речных. Такое поведение можно объяснить как упомянутой выше гипотезой «Лучше хоть что-то», так и гипотезой «Повышения плодовитости» (Fecundity enhancement) (Lyon, Eadie, 2008). Согласно последней, утка, откладывая несколько первых яиц в чужие гнезда, а уже потом в собственную кладку, повышает свою приспособленность. В характере межгодовой динамики уровня гнездового паразитизма у красноголового нырка и хохлатой чернети имеются различия. У красноголового нырка высокий общий уровень паразитизма наблюдался при любой численности. В годы, когда доля его гнезд на озере снижалась, повышался уровень межвидового паразитизма, и наоборот. У хохлатой чернети при снижении доли ее гнезд в утином сообществе не происходило повышения уровня межвидового паразитизма по сравнению с уровнем внутривидового. Вероятно, эти различия обусловлены сроками размножения этих видов. Красноголовый нырок – рано гнездящийся вид. У него сезон откладки яиц перекрывается с размножением и рано-, и поздногнездящихся видов. Поэтому он имеет возможность в течение более длительного периода использовать гнезда других видов. Хохлатая чернеть – поздноразмножающийся вид. Сроки ее гнездования практически совпадают с размножением серой утки. Это и обусловило высокую частоту паразитизма хохлатой чернети в гнездах этого вида.
Заключение
Все рассмотренные нами виды уток принимают двойное участие в явлении гнездового паразитизма. Во-первых, в их гнезда подкладывают яйца как конспецифичные особи, так и самки других видов. Во-вторых, каждый вид участвует в качестве паразита как во внутривидовых смешанных кладках, так и в межвидовых. Преобладающие по численности на озере кряквы, красноголовые нырки и хохлатые чернети достоверно чаще подкладывали яйца в гнезда своего вида. Немногочисленные виды (серая утка, красноносый нырок) – в гнезда других видов уток. Тем не менее, с учетом процентного соотношения числа утиных гнезд на озере, все виды предпочитали гнезда конспецифичных особей.
Уровень гнездового паразитизма нырковых уток выше, чем у речных. Высокий общий уровень паразитизма красноголового нырка не зависел от численности этого вида на озере. При снижении доли его гнезд он в равной мере паразитировал на кладках других видов уток. При увеличении – на кладках своего вида. Не меньшую склонность к гнездовому паразитизму проявил красноносый нырок. У хохлатой чернети общий уровень гнездового паразитизма был в среднем ниже, чем у красноголового и красноносого нырка. С увеличением на озере числа гнезд хохлатой чернети он достоверно увеличивался, главным образом за счет паразитических яиц во внутривидовых смешанных кладках.
Из речных уток, ежегодно гнездящихся на озере, серая утка по уровню гнездового паразитизма была на первом месте. Несмотря на низкую численность этой утки, при увеличении доли ее гнезд в утином сообществе достоверно увеличивалась доля яиц, подложенных в кладки конспецифичных особей. В условиях оз. Кротово кряква менее других видов была склонна к паразитизму. Однако и у нее прослеживалась тенденция повышения уровня и внутривидового, и межвидового паразитизма с увеличением числа гнезд.
Библиография
Волгин М. В., Сипко Л. Л. Физико-географическая и гидрохимическая характеристика Карасукских озер // Опыт комплексного изучения и использования Карасукских озер / Под ред. Г. М. Кривощекова. Новосибирск: Наука, 1982. С. 5–54.
Меднис А. А. Биология гнездования уток на озере Энгурес // Экология водоплавающих птиц Латвии / Под ред. Х. А. Михельсона. Рига: Зинатне, 1968. С. 85–108.
Меднис А. А., Блум П. Н. Отлов насиживающих уток и их птенцов // Кольцевание в изучении миграций птиц фауны СССР / Под ред. В. Д. Ильичева. М.: Наука, 1976. С. 157–167.
Михантьев А. И. Некоторые эколого-этологические механизмы регуляции численности утиных в природе и возможности их использования в биотехнии // Биотехния. Теоретические основы и практические работы в Сибири / Под ред. С. С. Фолитарека. Новосибирск: Наука, 1980. С. 146–190.
Михантьев А. И., Селиванова М. А. Вариации величины кладки у уток // Сибирский экологический журнал. 2008. Т. 15. № 1. С. 187–194.
Михантьев А. И., Селиванова М. А. Водоплавающие и околоводные птицы // Биоразнообразие Карасукско-Бурлинского региона (Западная Сибирь) / Под ред. Ю. С. Равкина. Новосибирск: Изд-во СО РАН, 2010. С. 215–233.
Нумеров А. Д. Межвидовой и внутривидовой гнездовой паразитизм у птиц . Воронеж: ФГУП ИПФ Воронеж, 2003. 517 с.
Пыжьянов С. В., Березовская А. О. Гнездовой паразитизм длинноносого крохаля Mergus serrator L., 1758 // Байкальский зоологический журнал. 2010. № 2 (5). 69–74.
Яновский А. П., Богдановская Н. И. Успешность размножения уток на оз. Мензелинское (Западная Сибирь) // Размещение и численность позвоночных животных Сибири. Новосибирск: Наука, 1982. С. 154–162.
Amat J. A. Parasitic Laying in Red-Crested Pochard Netta rufina Nests // Ornis Scandinavica. 1993. Vol. 24. № 3. P. 65–70. doi:10.2307/3676412.
Beauchamp G. The relationship between intra- and interspecific brood amalgamation in waterfowl // The Condor. 1998. Vol. 100. P. 153–162. doi:10.2307/1369908.
Deng Q.-X., Wang H.-T., Yao D., Wang X.-Y., E M.-J., Wang T., Gao W. Conspecific brood parasitism and nesting biology of Mandarin ducks (Aix galericulata) in Northeastern China // The Wilson Journal of Ornithology. 2011. Vol. 123. № 3. P. 479–485. http://www.jstor.org/stable/23033553.
Duebbert H. F., Lokemoen J. T., Sharp D. E. Concentrated Nesting of Mallards and Gadwalls on Miller Lake Island, North Dakota // The Journal of Wildlife Management. 1983. Vol. 47. № 3. P. 729–740. doi:10.2307/3808608.
Dugger B. D., Blums P. Effect of conspecific brood parasitism on host fitness for Tufted Duck and Common Pochard // The Auk. 2001. Vol. 118. № 3. P. 717–726. doi:10.2307/4089933.
Eadie J. M., Sherman P. W., Semel B. Conspecific Brood Parasitism, Population Dynamics, and the Conservation of Cavity Nesting Birds // Behavioral Ecology and Conservation Biology / T. Caro (ed.). Oxford: Oxford University Press, 1998. P. 306–340.
Geffen E., Yom-Tov Y. Factors affecting the rates of intraspecific nest parasitism among Anseriformes and Galliformes // Animal Behaviour. 2001. Vol. 62. № 6. P. 1027–1038. doi:10.1006/anbe.2001.1855.
Hines J. E., Mitchell G. J. Parasitic laying in nests of gadwalls // Canadian Journal of Zoology. 1984. Vol. 62. P. 627–630. doi:10.1139/z84-092
Kreisinger J., Munclinger P., Javůrková V., Albrecht T. Analysis of extra-pair paternity and conspecific brood parasitism in mallards Anas platyrhynchos using non-invasive techniques // Journal of Avian Biology. 2010. Vol. 41. № 5. P. 551–557. doi:10.1111/j.1600-048X.2010.05002.x.
Lebedeva, N. V., Markitan, L. V. Problems of population dynamics of the White-Eyed Pochard (Aythya nyroca Guld., 1770) in the Eastern Sea of Azov Region // Russian Journal of Ecology. 2001. Vol. 32. № 6. P. 459–465. https://doi.org/10.1023/A:1012538302166.
Lokemoen J. T. Brood Parasitism among Waterfowl Nesting on Islands and Peninsulas in North Dakota // The Condor. 1991. Vol. 93. P. 340–345. doi:10.2307/1368949.
Lyon B. E., Eadie J. M. Mode of development and interspecific avian brood parasitism // Behavioral Ecology. 1991. Vol. 2. № 4. P. 309–318. doi:10.1093/beheco/2.4.309.
Lyon B. E., Eadie J. M. Conspecific brood parasitism in birds: a life-history perspective // Annual Review of Ecology, Evolution and Systematics. 2008. Vol. 39. P. 343–363. doi:10.1146/annurev.ecolsys.39.110707.173354.
Musil P., Neužilova Š. Long-term changes in duck inter-specific nest parasitism in South Bohemia, Czech Republic // Wildfowl. 2009. Special Issue 2. P. 176–183.
Peters J. L., Brewer G. L., Bowe L. M. Extrapair Paternity and Breeding Synchrony in Gadwalls (Anas strepera) in North Dakota // Auk. 2003. Vol. 120. № 3. P. 883–888. doi:10.2307/4090119.
Petrželková A., Klvaňa P., Albrecht T., Hořák D. Conspecific brood parasitism and host clutch size in Common Pochards Aythya ferina // Acta Ornithologica. 2013. Vol. 48. № 1. P. 103–108. doi:10.3161/000164513X670052.
Pöysä H., Eadie J. M., Lyon B. E. Conspecific brood parasitism in waterfowl and cues parasites use // Wildfowl. 2014. Special Issue 4. P. 192–219.
Roy C., Eadie J. M., Schauber E. M., Odell N. S., Berg E. C., Moore T. Public information and conspecific nest parasitism in wood ducks: does nest density influence quality of information? // Animal Behaviour. 2009. Vol. 77 P. 1367–1373. http://doi.org/10.1016/j.anbehav.2009.02.017.
Sayler R. D. Ecology and evolution of brood parasitism in waterfowl // Ecology and Management of Breeding Waterfowl / B. D. J. Batt, A. D. Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec, G. L. Krapu (eds.). Minneapolis: University of Minnesota Press, 1992. P. 290–322.
Sorenson M. D. Comment: Why is conspecific nest parasitism more frequent in waterfowl than in other birds? // Canadian Journal of Zoology. 1992. Vol. 70. № 9. P. 1856–1858. doi:10.1139/z92-253.
Šťovíček O., Kreisinger J., Javůrková V., Albrecht T. High rates of conspecific brood parasitism revealed by microsatellite analysis in a diving duck, the common pochard Aythya ferina // Journal of Avian Biology. 2013. Vol. 44. № 4. P. 369–375. doi:10.1111/j.1600-048X.2013.00074.x.
Sukhanova O.V. Nesting Ecology of the Tufted Duck (Aythya fuligula) and the Pochard (Aythya ferina) in Central Russia // Anatidae 2000: An International Conference on the Conservation, Habitat Management and Wise Use of Ducks, Geese and Swans. Gibier Faune Sauvage Game and Wildlife. 1996. Vol. 13. № 2. P. 709–722.
Titman R. D., Lowther J. K. The breeding behavior of a crowded population of mallards // Canadian Journal of Zoology. 1975. Vol. 53. № 9. P. 1270–1283. doi:10.1139/z75-152.
Waldeck P., Kilpi M., Öst M., Andersson M. Brood Parasitism in a Population of Common Eider (Somateria mollissima) // Behaviour. 2004. Vol. 141. P. 725–739. http://www.jstor.org/stable/4536162.
Westerskov K. Methods for Determining the Age of Game Bird Eggs // The Journal of Wildlife Management. 1950. Vol. 14. № 1. P. 56–67. doi:10.2307/3795978.
Yom-Tov Y. An updated list and some comments on the occurrence of intraspecific nest parasitism in birds // Ibis. 2001. Vol. 143. № 1. P. 133–143. doi:10.1111/j.1474-919X.2001.tb04177.x.
Благодарности
Мы благодарны за помощь студентам-дипломникам и аспирантам, участвовавшим в разные годы в сборе полевого материала. Также мы признательны покойному д. б. н. С. С. Фолитареку за его советы на первых этапах наших исследований. Мы благодарны зав. биостанцией ИСиЭЖ СОРАН В. А. Шило, все эти годы оказывавшему нам всяческое содействие в организации быта во время экспедиционных работ. Исследование частично финансировалось по проекту № VI.51.1.8 программы ФНИ государственных академий наук на 2013–2020 гг., а также по проекту РФФИ № 15-29-02479-офи-м.

 © 2011 - 2024
© 2011 - 2024