



Введение
Живорождение – важнейшая стратегия выживания рептилий на Севере. Сама возможность вынашивания потомства открывает путь для следующей адаптации – продления беременности до момента оптимального развития молоди. Этот момент в конце короткого (90 дней) северного лета может и не наступить. Как теперь известно, в ответ на изменчивость погоды популяции гадюковых змей выработали важную адаптацию – беременные самки асписовой и обыкновенной гадюк зимуют с эмбрионами. Это явление известно для Франции, Казахстана, Швеции (Saint-Girons, Duguy, 1973; Бердибаева, 1981; Madsen, 1989) и для Карелии (Коросов, 2000). Вопрос состоит в том, насколько широко оно распространено в популяции гадюк Карелии?
Материалы
Плодовитость изучалась в июле, августе и сентябре 1998 г. (учтено 252 экз.), в мае2007 г. (71 экз.) на островах Кижского архипелага (Заонежье, Онежское озеро), в том числе на о. Кижи.
Методы
Плодовитость оценивалась по числу эмбрионов, обнаруженных путем пальпирования у живых самок гадюки. Точность прижизненной оценки плодовитости контролировалась по трупному материалу (34 экз.) (пищевой комок, который располагается много выше яйцеводов, мы не могли принять за эмбрионов). В начале августа (за месяц до рождения) эмбрионы свернуты в относительно плотную спираль и хорошо прощупываются. Пальпирование довольно грубая процедура, которая может привести к травмам эмбрионов (Кудрявцев, 1991), поэтому на о. Кижи она проводилась только в течение двух сезонов во избежание нарушения динамики размножения гадюк.
Результаты
Вскрытие 4 взрослых самок обыкновенной гадюки, закончивших период спаривания весной 1998 г. (20 мая), показали, что в одной из самок (размеры: L. t. = 620 мм, L. c. = 85 мм, W = 165 г) находились пять хорошо развитых эмбрионов самок (2 экз.) и самцов (2 экз.), заключенных в тонкую прозрачную оболочку, а также еще один самец, заключенный в плотную толстую гиперимированную оболочку (рис. 1).

Рис. 1. Вскрытая самка обыкновенной гадюки с эмбрионами (А, Б) и зрелыми яйцами (В)
Fig. 1. Female common viper with embryos (A, Б) and mature eggs (B)
Размеры эмбрионов близки к размерам августовских новорожденных: у самок L. t. = 152 мм, L. c. = 19.5 мм, W = 3.25 г, у самцов – L. t. = 146.5 мм, L. c. = 25 мм, W = 3.4 г. (в среднем). Тела четырех первых эмбрионов были свободно уложены по спирали, а последний был плотно упакован (рис. 1, Б). По всей видимости, этот эмбрион находился на ранней стадии резорбции, а 4 других эмбриона должны были вскоре родиться. Важно отметить, что только этот эмбрион пальпировался как крупный плотный комок, остальные эмбрионы были неотличимы по консистенции от яйцеводов, наполненных зрелыми яйцами, у других самок. Помимо эмбрионов, яйцеводы гадюки содержали 8 крупных зрелых яиц (размеры от 33 х 11 до 35 х 12 мм) (рис. 1, В). Итак, эта самка не только сохранила беременность с прошлого года, но могла бы размножаться и в текущем, т. е. становиться беременной каждый год!
Ключевое значение в этом играет, на наш взгляд, хорошая упитанность крупной немолодой змеи. Как показывают наши данные мечения гадюк в течение одного сезона, потери массы тела при родах у самок разного размера составляют от 20 до 60 % (рис. 2, 1, 3). К сожалению, в эту выборку не попали змеи тяжелее 160 г. Если включить в выборку большее число особей (охватив более длительный период между повторными отловами – 1 и 2 года), то оказывается, что самые большие сезонные потери массы самых крупных особей достигают всего 5 % и они много меньше, чем потери массы мелких (молодых) особей (до 60 %).
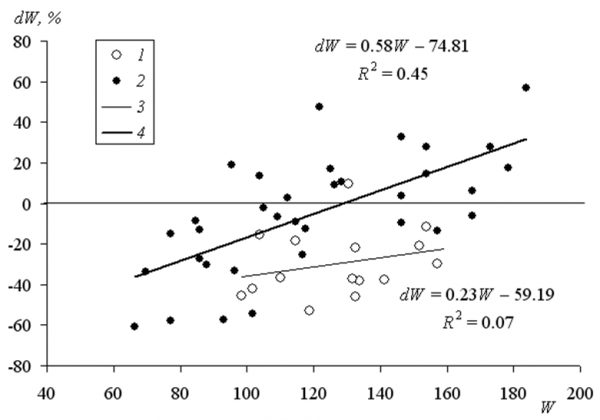
Рис. 2. Изменение массы тела (dW = Wi+1 – Wi, %) самок гадюки между двумя отловами:
1 – лето – осень, 2 – веснаi+1 – веснаi, 3 – тренд для 1, 4 – тренд для 2; R2 – коэффициенты детерминации трендов
Fig. 2. Change in body weight (dW = W i+1 – W i, %) of viper females between two catches:
1 – summer – autumn, 2 – springi+1 – springi, 3 – the trend for the 1,
4 – the trend for 2; R2 – coefficients of determination of trends
Различие между мелкими (молодыми) и крупными (старшими) змеями состоит еще и в том, что молодые гадюки в гораздо меньшей степени способны быстро восстановить полученный дефицит массы, чем крупные старшие особи. В зависимости от возраста (и размера) на восстановление массы тела, необходимой для следующего размножения, гадюкам требуется до трех лет (Volsøe, 1944; Viitanen, 1967; Biella, 1979; Shane, Seigal, 1989). Ориентируясь на обнаруженные соотношения (рис. 1), можно констатировать, что самки, весящие больше160 г, после родов существенно не теряют в массе. Видимо, эта категория самок может размножаться каждый год и при неблагоприятных условиях уходить на зимовку с эмбрионами. Судя по размерам тела (Коросов, 2010), возраст у этих животных должен составлять не менее 10 лет.
Обсуждение
В рамках исследования популяционной репродукции важно хотя бы приблизительно оценить долю самок, зимующих с эмбрионами или размножающихся ежегодно. Поскольку обыкновенная гадюка – редкое животное, ее невозможно добывать и препарировать в большом количестве. Выполнить поставленную задачу позволит, на наш взгляд, анализ распределения самок по массе тела и регрессия массы тела на линейные размеры. Нами показано, что масса самок с эмбрионами к концу беременности увеличивается (Коросов, 2010). Это значит, что весной беременные самки будут самыми тяжелыми среди голодных животных этого размерного класса. Наши материалы по массе самок гадюки, отловленных весной 1991–2009 гг., показывают, что в популяции существует очень небольшое число особенно крупных особей с длиной тела более62 сми массой – более160 г. Среди 1450 изученных самок их было всего 18 экз., или 1.2 % (рис. 3).
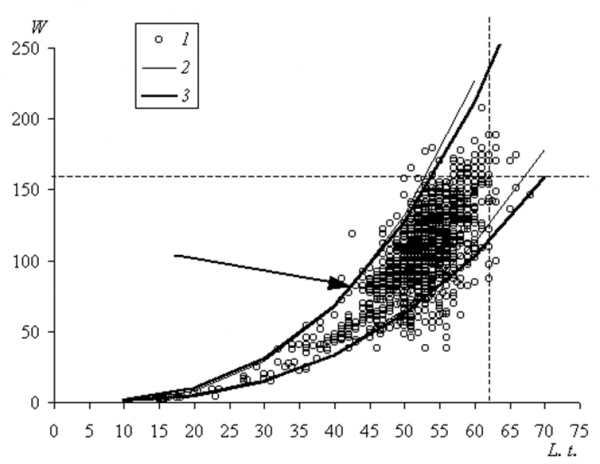
Рис. 3. Доверительные зоны для вариант, построенные вокруг линий регрессии массы тела (W, г) по длине тела (L. t., см) самок гадюки: 1 – исходные денные, 2 – экстраполяция доверительной зоны регрессии, построенной для особей с длиной тела менее 45 см, 3 – доверительная зона регрессии для всех особей; стрелкой указано сгущение точек, соответствующих впервые беременным самкам
Fig. 3. Confidence zones for the variants, built around the regression lines of body weight (W, g) on body length (L. t., cm) of female vipers: 1 – initial data, 2 – extrapolation of the confidence zone regression, built for individuals with a body length less than 45 cm, and 3 – confidence zone regression for all individuals, the arrow indicates the concentration of points corresponding to the first pregnant females
Возможно, особи меньшего размера также могут обладать способностью к частому размножению, но их доля вряд ли может превысить 3 % (доля самок тяжелее 160 гсоставляет 3.7 %, а длиннее62 см– 1.9 %). Итак, есть основания утверждать, что доля самок, зимующих с эмбрионами ( и размножающихся каждый год) составляет менее 3 % (крупные особи в возрасте 10–15 лет).
Один из возможных вариантов выявления таких особей состоит в том, чтобы учитывать «аномально» тяжелых для своих размеров «пожилых» особей. Если ежегодное размножение и зимовка с эмбрионами связанны только с индивидуальными особенностями и не является обычным явлением у самых крупных растущих самок, то такие особи должны находиться за пределами верхней границы доверительного интервала для беременных самок (принят уровень доверительной вероятности P = 0.9).
С этой целью рассчитали два уравнения степенной регрессии массы тела по размерам и их доверительные зоны (для каждого p < 0.05). Первая кривая L.t. = 0.0008W 2.98 построена по данным для молодых неполовозрелых особей (с размерами тела меньше 45 см) (рис. 3, 2); смысл ее построения состоял в том, чтобы выяснить, в какой мере пропорции тела, заложенные в молодости, сохраняются и во взрослом состоянии. Вторая кривая L.t. = 0.0007W 3.00 построена для особей всех возрастов и характеризует весь эмпирический материал (рис. 3, 3). Можно было ожидать, что доверительная зона регрессии, построенной по данным для молодых, не претерпевших связанные с беременностью рост и падение массы, будет более узкой, чем зона для беременных, однако обе границы зон практически совпали. Варьирование массы тела, связанное с беременность, как бы «вписано» в видовую норму и детерминировано уже на ранних этапах роста самок.
В нашей выборке за пределами верхней границы находится группа из 15 аномально тяжелых особей (0.8 %). Часть из них, видимо, незадолго перед отловом питалась и (случайно) не была идентифицирована и не отделена от остальных особей; это относится к молодым особям длиной менее45 см(3 точки).
Вторая группа из 5 точек соответствует средневозрастным взрослым особям, превышение массы которых над максимальными значениями очень невелико (одна из возможных причин – отлов особей через 4-5 дней после охоты, когда добыча уже почти переварена и не пальпируется, но остатки которой еще не превратились в экскреты). Однако эта прибавка (5–11 г) не может играть существенной роли при общих потерях при родах в50 ги, следовательно, способствовать ежегодному размножению. Наиболее интересна третья группа из 4–6 особей, масса которых на 15–20 г выше максимальной границы. Они вполне могут быть отнесены к группе ежегодно размножающихся (или зимующих с эмбрионами); в уловах их доля составляет 0.4 %.
Нижняя доверительная граница соотношения масса/длина, построенная для всех особей, также практически совпадает с границей, построенной для молодых особей. Она отделяет тех особей, у которых при родах происходит существенная потеря массы, в определенной мере аномальная для обычных пропорций самок. Особенно сильно эта потеря выражена у змей с длиной тела меньше58 см; при визуальном осмотре их состояние кажется просто критическим (прощупывается скелет, кожа образует продольные складки). Есть основания считать, что такие животные не могут пережить зиму (Madsen, 1978).
Судя по нашим данным, самки гадюки, способные к зимовке с эмбрионами и размножению каждый год, обычно не представляют собой какой-либо аномалии (соответствующие им точки находятся в пределах доверительной зоны), это просто «пожилые» и крупные животные. Однако в популяции есть и менее крупные лидеры по упитанности, видимо, способные легко перенести потерю массы при родах. Общая доля все этих животных составляет около 2–3% от числа половозрелых самок.
На наш взгляд (судя по географии находок: северная Европа – южная Карелия – северный Казахстан) зимовка самок обыкновенной гадюки с эмбрионами – не только потенциально возможное, но и широко распространенное явление для условий сокращенного теплого периода. Возникает вопрос, почему имеются лишь единичные его описания? Первая причина состоит в том, что в южной части ареала гадюки обычно рождают молодых задолго до наступления неблагоприятных погодных условий. Во-вторых, старые крупные особи – достаточная редкость для популяции гадюки с предельным возрастом жизни 11-13 лет. Наконец, обнаружение факта зимовки гадюки с эмбрионами затруднено, поскольку при пальпировании на последних стадия развития (весной) эмбрионы не прощупываются (к тому же их можно спутать со зрелыми яйцами), а случаи вскрытия животных описанной группы немногочисленны (и вряд ли оправданы в большом объеме).
Учитывая способность размножаться каждый год, следует пересчитать роль описанной группы самок в популяционной репродукции. Поскольку из числа половозрелых гадюк только треть размножается ежегодно (Коросов, 2010), доля молодых, рожденных группой крупных самок, может доходить до 5–9% ото всех новорожденных. Для территории южной Карелии эта величина не является существенной. Однако в отдельные годы и на севере карело-кольского края, где погодные условия, благоприятные для нормального размножения, могут отсутствовать несколько лет подряд (Коросов, Ивантер, 2003), вклад крупных самок, зимующих с эмбрионами, может оказаться определяющим для судьбы локальной популяции.
Заключение
Наши исследования населения обыкновенной гадюки в южной части республики Карелия показали, что доля крупных самок способных к зимовки с эмбрионами и, види-мо, к ежегодному размножению, составляет менее 3% от половозрелых самок. Роль этих особей в репродукции местных популяций невелика.
Библиография
Бердибаева Ж. Ш. К биологии обыкновенной гадюки // Фауна и экология животных Ка-захстана. Алма-Ата, 1981. С. 31–33.
Коросов А. В. Рептилии Карело-Кольского края // Биогеография Карелии: Тр. КНЦ РАН. Петрозаводск, 2000. Вып. 2. С. 96–102.
Коросов А. В. Экология обыкновенной гадюк (Vipera berus L.) на Севере (факты и моде-ли). Петрозаводк: Изд-во ПетрГУ, 2010. 264 с.
Коросов А. В., Ивантер Э. В. Изменчивость фенологических явлений в жизни гадюки в условиях Карелии // Змеи Восточной Европы. Тольятти, 2003. С. 34.
Кудрявцев С. В., Фролов В. Е., Королев А. В. Террариум и его обитатели. М., 1991. 296 с. Biella H.-J. Untersuchungen zur Fortpflanzungsbiologie der Kreuzotter (Vipera berus L.) (Reptilia, Serpentes, Vipiridae) // Zool. Abh. Staatl. Mus. Tierk. Dresden. 1979. Vol. 36. P. 117–125.
Madsen Th. Reproducrive success, mortallity and sexual size dimorphism in the adder, Vipera berus, L. // Holarct. Ecol., 1978. 11. P.77-80.
Madsen Th. Female adder in southern Sweden recorded giving birth in spring // Amphibia Reptilia. 1989. Vol. 10. № 1. P. 88–89.
Saint-Girons H., Duguy R. Un exemple de predation atypique et de gestation hibernale chez Vipera aspis (L.) // Ann. Soc. Sci. natur. Charente-Naritime. 1973. Vol. 5. № 5. P. 379–381.
Viitanen P. Hidernation and seasonal movements of the viper, Vipera berus ( L.), in southern Finland // Ann. Zool. Fennici. Helsinki. 1967. Vol. 4. № 4. P. 472–546.
Volsøe H. Structure and seasonal variation of the male reproductive organs of Vipera berus ( L.). Copenhagen, 1944. 157 p. / Spolia zoologica Musei heuniensis.Surifter udg Uviv Zoologiske museum Kobenavn. 5.

 © 2011 - 2026
© 2011 - 2026