



Введение
В некрофильном комплексе жесткокрылые представлены достаточно большим числом видов. Например, в Чехии на трупах животных выявлено 145 видов из 22 семейств (Kočárek, 2003). В зоне средней тайги Карелии число некрофильных видов также высоко (Лябзина, 2011). На трупе жуки присутствуют всегда, и на протяжении всего процесса разложения численность их высока. Трофические взаимосвязи с трупом у жуков разнообразны. Встречаются некрофаги, кератофаги, кожееды, хищники, паразитоиды, сапрофаги, детритофаги, мицетофаги и др. Большая часть из них относятся к факультативным некробионтам и кроме трупа встречаются на других эфемерных органических субстратах (навоз, грибы). В этой связи возникает вопрос: оказывают ли влияние на некробионтов факторы внешней среды, различающиеся в разных местообитаниях, или же определяющим является только наличие трупного материала?
Целью работы являлось изучение биотопической специфики фауны некробионтов с помощью разнообразных математических средств обработки данных по встречаемости этих животных на трупах. Для придания работе большей прозрачности ограничились анализом учетов в трех контрастных типах биотопов – сосняк, березняк, луг.
Материалы
Исследования проводили в весенне-осенние периоды 2000–2012 гг. в окрестностях биологической станции ПетрГУ (с. Кончезеро, Кондопожский р-н) (34°00'E 62°06'N). В сообщении приводятся данные по трем типам биотопов: сосняк чернично-зеленомошный, березняк разнотравный и влажно-разнотравный луг. Выбранные биотопы типичны для средней подзоны тайги (Раменская, 1958; Волков, 2008). Жуков-мертвоедов собирали с трупных приманок, выложенных в разные годы, – по 9 в каждом биотопе (всего 27). Всего было собрано более 4 тысяч экземпляров жуков, относящихся к 85 видам.
Исходные данные для статистических расчетов представляют собой таблицу, составленную из 28 столбцов (название вида и 27 приманок) и 85 строк (по одной для каждого из 85 видов), в которую занесены значения количества жуков каждого вида, отловленных во время разового учета на каждой трупной приманке.
Методы
В работе применяли общепринятую методику по сбору фауны некробионтов. Трупы выкладывали на поверхность почвы либо помещали в почвенные ловушки (банки диаметром входного отверстия 7.5 см и объемом 500 мл). Все приманки раскладывали на расстояние более 100 м друг от друга. Приманками служили трупы беспозвоночных и позвоночных животных массой от 30 г до 4 кг (навеска трупной массы брюхоногих моллюсков, дождевых червей или отдельные тушки плотвы, гадюки, сороки, вороны, чайки сизой, полевки рыжей, крота, белки, куницы, кошки). В каждом исследованном биотопе раскладывали по девять приманок. Сбор насекомых производился с самого трупа и с подстилки под ним. Отлов жуков осуществляли в течение всего периода разложения.
В разных процедурах расчета использовались разные значения оценок обилия животных. Для расчета индексов видового богатства относительное обилие видов оценивали по 4-балльной логарифмической шкале (Песенко, 1982). Сначала общее число жуков всех видов, встреченных на каждом трупе, усредняли по всем трупам для каждого биотопа. Затем полученные величины общего обилия N преобразовывали в баллы. Балльная шкала представлена последовательно увеличивающимися значениями границ для серии классовых интервалов, которые рассчитывались по формулам:
Верхняя граница класса на шаге a:
N a/k, (a = 1, 2, …, k), где N – объем всей выборки, a – шаг шкалы, k – число классов.
Формула длины интервала а-го шага шкалы:
ba = Na/k – N(a-1)/k.
Балл обилия i-го вида с числом особей в выборке ni является целым числом, заключенным в интервале значений:
K logN ni < a < (k logN ni + 1).
Вычисленные значения границ классового интервала при сборе 4499 особей жесткокрылых-некробионтов при четырех баллах обилия представлены в таблице 1.
Таблица 1. Четырехбалльная шкала относительного обилия жесткокрылых-некробионтов
Table 1. Four-point scale relative abundance beatles collected from carcasses of animals
| Балл, a | Вид (по обилию) | Граница классового интервала | |
| нижняя | верхняя | ||
| 1 | Редкий (единично) | 1 | 8 |
| 2 | Обычный | 9 | 67 |
| 3 | Многочисленный | 68 | 549 |
| 4 | Массовый | 550 | 4499 |
Разнообразие жесткокрылых-некробионтов оценивали с помощью часто используемых индексов, характеризующих сообщества в биоценозе (Мэгарран, 1992; Коросов, 2007). Индекс Шеннона (Н2) определяет видовое богатство: чем больше его значения, тем выше видовое разнообразие сообщества. Индекс Пиелу (Е) характеризует выравненность видов в сообществе. Величина индекса Пиелу изменяется от 0 до 1, при единице сообщество характеризуется равным обилием всех видов. Индекс доминирования Бергера – Паркера (d) выражает относительную значимость наиболее обильного вида. Увеличение индекса Бергера – Паркера означает уменьшение разнообразия и увеличение степени доминирования одного вида. Число доминирующих видов («эвивалентного сообщества») определяли по формуле sH = 2H (Песенко, 1982; Коросов, 2007).
Для всех прочих расчетов использовали значения количества жуков-мертвоедов, встреченных на каждой трупной приманке. Показатели обилия (ni) предварительно были нормированы (автошкалированы) с целью устранить влияние размера трупного объекта на число встреченных мертвоедов. Сначала для жуков всех видов на одном трупе рассчитывали среднее значение численности M и стандартное отклонение S. Затем для каждого вида на данном трупе определяли величину ni’ = (ni – M) / S.
Сопоставление нормированных оценок обилия жуков позволило выявить виды как с очень высокими показателями, так и близкими к нулю. Большой разрыв между значениями может привести к появлению очень высоких корреляций между столбцами исходной матрицы и, как следствие, к искажению результатов компонентного анализа. Исходя из этих соображений была составлена дополнительная таблица, в которую вошли виды со средним (по всем спискам) нормированным значением обилия больше –0.3 и меньше 2; исходное число видов 85 сократилось до 29. Так получили таблицу нормированных значений встречаемости 29 видов на 27 приманках.
Эти данные были использованы для выполнения компонентного анализа (Коросов, 1996, 2007) и для статистической оценки значимости различий показателей обилия некробионтов в разных биотопах с помощью бутстрепа (Шитиков, 2012).
Компонентный анализ изучает корреляцию между характеристиками (столбцами матрицы), в роли которых выступали трупы животных. Благодаря процедуре разложения корреляций на агенты влияния (факторы) на их фоне удается выявить группы показателей, обладающих более тесными связями, т. е. сходно реагирующих на выявленные факторы. Эта процедура использована нами для иллюстрации различия видового состава некробионтов в разных биотопах.
Бутстреп-анализ позволяет оценить значимость различия между выборками безотносительно к законам распределения численных характеристик; в нашем случае – проверить нулевую гипотезу о равенстве показателей обилия фауны мертвоедов трех биотопов. Выполняли попарное сравнение биотопов. В качестве меры различий между двумя группами трупов (биотопами) взяли евклидово расстояние между средними значениями нормированной численности животных: d12 = (Σ(n1s’– n2s’)2)½, где n1s’, n2s’ – нормированные значения обилия вида s в биотопах 1 и 2. Таким образом, процедура расчета исходных расстояний dи состоит из двух этапов. Сначала рассчитываются три столбца средней нормированной численности всех видов nis’ – по одному для каждого биотопа. Затем для каждой пары столбцов рассчитываются три значения расстояний: dи12, dи13 и dи23.
Технология бутстрепа состоит в следующем. Из общего набора данных (27 столбцов) случайным образом формировались две группы из 9 векторов оценок численности мертвоедов с разных приманок. Для каждой группы вычислялись векторы средней оценки численности жуков, между которыми вычислялось «случайное» евклидово расстояние dс. Процесс случайного извлечения выборок выполнялся по 3000 раз. Полученные наборы случайных оценок евклидовых расстояний dс использовались для построения их распределения и расчета квантилей для p = 95 %. Поскольку метрика d положительна и симметрична, в качестве критической границы брали только правую (квантиль dс0.95). Далее оценка расстояния между выборками из разных биотопов dи сравнивалась со значением квантили dс0.95, полученным бутстрепом. Если величина dи превышала значение квантили, различие между выборками считали значимым и рассматривали как доказательство реальности влияния биотопов на структуру населения. Все расчеты выполнены в среде пакетов Excel и R.
Результаты
На трупах животных в трех исследуемых биотопах выявлено 85 видов жесткокрылых из 14 семейств (табл. 2).
Таблица 2. Видовой состав и балл обилия жесткокрылых-некробионтов
Table 2. Species composition and abundance score of Coleoptera species collected during the study of carcass decomposition in Karelia and their point scale relative abundance
| Сосняк | Березняк | Луг | |
| Сем. Carabidae | |||
| Pterostichus niger (Schaller, 1783) | 0 | 1 | 1 |
| Сем. Hydrophilidae | |||
| Cryptopleurum crenatum (Kugelan, 1794) | 0 | 1 | 0 |
| C. minutum (Fabricius, 1775) | 2 | 1 | 2 |
| Cyceron impressus (Sturm, 1807) | 3 | 2 | 2 |
| C. lateralis (Marsham, 1802) | 3 | 2 | 2 |
| C. unipunctatus (Linnaeus, 1758) | 1 | 1 | 1 |
| Megasternum concinnum (Marsham, 1802) | 0 | 1 | 0 |
| Сем. Histeridae | |||
| Gnathoncus nanus (Scriba, 1790) | 1 | 0 | 0 |
| Hister unicolor (Linnaeus, 1758) | 0 | 1 | 1 |
| Margarinotus cadaverinus (Hoffmann, 1803) | 3 | 3 | 3 |
| Saprinus aeneus (Fabricius, 1775) | 0 | 0 | 1 |
| S. immundus (Gyllenhal, 1808) | 0 | 0 | 1 |
| S. planiusculus (Motschulsky, 1849) | 0 | 0 | 2 |
| S. semistriatus (Scriba, 1790) | 2 | 2 | 3 |
| Сем. Leiodidae | |||
| Apocatops nigrita (Fabricius, 1792) | 0 | 1 | 2 |
| Sciodrepoides watsoni (Spence, 1815) | 3 | 3 | 2 |
| S. fumatus (Spence, 1815) | 3 | 3 | 3 |
| Сем. Silphidae | |||
| Necrodes littoralis (Linnaeus, 1758) | 2 | 2 | 2 |
| Nicrophorus invetstigator (Zetterstedt, 1824) | 2 | 2 | 2 |
| N. vespillo (Linnaeus, 1758) | 1 | 1 | 2 |
| N. vespilloides (Herbst, 1783) | 3 | 3 | 2 |
| Oiceoptoma thoracicum (Linnaeus, 1758) | 3 | 3 | 0 |
| Silpha carinata (Herbst, 1783) | 0 | 0 | 0 |
| S. obscura (Linnaeus, 1758) | 0 | 0 | 1 |
| S. tristis (Illiger, 1798) | 0 | 0 | 1 |
| Thanatophilus dispar (Herbst, 1793) | 0 | 0 | 1 |
| Th. rugosus (Linnaeus, 1758) | 0 | 0 | 2 |
| Th. sinuatus (Fabricius, 1775) | 0 | 0 | 4 |
| Сем. Staphylinidae | |||
| Aleochara curtula (Goeze, 1777) | 4 | 4 | 4 |
| A. bipustulata (Linnaeus, 1761) | 1 | 0 | 1 |
| A. brevipennis (Gravenhorst, 1806) | 1 | 0 | 0 |
| Atheta corvina (Thomson, 1856) | 0 | 2 | 0 |
| A. crassicornis (Fabricius, 1792) | 3 | 2 | 3 |
| A. boleticola (Sahl, 1876) | 1 | 0 | 0 |
| A. divisa (Märkel, 1844) | 0 | 2 | 1 |
| A. europaea (Likovský, 1984) | 0 | 0 | 1 |
| A. fungi (Gravenhorst, 1806) | 0 | 1 | 0 |
| A. gagatina (Baudi, 1848) | 0 | 0 | 1 |
| A. nigripes (Thomson, 1856) | 0 | 0 | 1 |
| A. paracrassicornis (Brundin, 1954) | 4 | 4 | 4 |
| A. pseudotenera (Cameron, 1933) | 0 | 1 | 0 |
| A. strandiella (Brundin, 1954) | 0 | 1 | 0 |
| A. sylvicola (Kraatz, 1856) | 1 | 0 | 0 |
| Gyrohypnus angustatus (Stephens, 1833) | 0 | 1 | 1 |
| G. punctulatus (Paykull, 1789) | 1 | 0 | 0 |
| G. fracticornis (Miiller, 1776) | 1 | 0 | 0 |
| Lathrobium sp. | 0 | 1 | 0 |
| Liogluta microptera (Thomson, 1867) | 0 | 0 | 0 |
| Creophilus maxillosus (Linnaeus, 1758) | 2 | 2 | 2 |
| Deliphrum tectum (Paykull, 1789) | 2 | 2 | 2 |
| Megarthrus denticollis (Beck, 1817) | 1 | 2 | 1 |
| Omalium rivulare (Paykull, 1789) | 1 | 2 | 2 |
| O. septentrionis (Thomson, 1857) | 2 | 2 | 0 |
| Ontholestes murinus (Linnaeus,1758) | 2 | 2 | 3 |
| O. tessulatus (Geoffroy, 1785) | 2 | 2 | 2 |
| Anotylus rugosus (Fabricius, 1775) | 2 | 2 | 1 |
| Proteinus brachypterus (Fabricius, 1792) | 2 | 2 | 3 |
| Philonthus addendus (Sharp, 1867) | 1 | 2 | 2 |
| Ph. fumarius (Gravenhorst, 1806) | 2 | 0 | 0 |
| Ph. laminatus (Creutzer, 1799) | 0 | 0 | 1 |
| Ph. marginatus (Stroem, 1768) | 0 | 1 | 0 |
| Ph. nitidus (Gravenhorst, 1802) | 0 | 0 | 2 |
| Ph. politus (Linnaeus 1758) | 2 | 1 | 0 |
| Ph. sanguinolentus (Gravenhorst, 1802) | 0 | 2 | 1 |
| Ph. succicola (Thomson, 1860) | 4 | 2 | 4 |
| Ph. varians (Paykull, 1789) | 2 | 0 | 2 |
| Platystethus arenarius (Geoffroy, 1785) | 0 | 0 | 1 |
| Proteinus brachypterus (Fabricius, 1792) | 1 | 2 | 1 |
| Rugilus rufipes (Germar, 1836) | 2 | 0 | 0 |
| Tachinus laticollis (Gravenhorst, 1802) | 2 | 4 | 1 |
| T. marginellus (Fabricius, 1792) | 3 | 2 | 1 |
| T. signatus (Gravenhorst, 1802) | 0 | 2 | 2 |
| T. proximus (Kraatz, 1855) | 0 | 1 | 0 |
| Сем. Ptiliidae | |||
| Acrotrichis parva (Rosskothen, 1935) | 2 | 0 | 0 |
| Сем. Aphodiinae | |||
| Aphodius rufipes (Linnaeus,1785) | 0 | 0 | 1 |
| Сем. Geotrupidae | |||
| Geotrupes stercorarius (Linnaeus, 1758) | 1 | 0 | 1 |
| Anoplotrupes stercorosus (Scriba, 1791) | 2 | 1 | 2 |
| Сем. Dermestidae | |||
| Dermestes ater (De Geer, 1774) | 1 | 0 | 0 |
| D. murinus (Linnaeus, 1758) | 1 | 0 | 0 |
| Сем. Cleridae | |||
| Necrobia violacea (Linnaeus, 1758) | 2 | 1 | 2 |
| Сем. Elateridae | |||
| Prosternon tessellatum (Linnaeus, 1758) | 0 | 2 | 0 |
| Сем. Nitidulidae | |||
| Omosita depressa (Linnaeus, 1758) | 2 | 1 | 2 |
| O. colon (Linnaeus, 1758) | 1 | 0 | 1 |
| Nitidula bipunctata (Linnaeus, 1758) | 0 | 2 | 1 |
| Сем. Cryptophagidae | |||
| Atomaria fuscata (Schönherr, 1808) | 0 | 1 | 0 |
На лугу отмечено наибольшее число жуков-мертвоедов – 58 видов, в березняке – 53, в сосняке – 49. Однако индекс разнообразия (Н) луговых биоценозов имеет наименьшее значение по сравнению с лесными биотопами (табл. 3). Это связано с большим числом встреченных здесь видов, которые относятся к первому баллу обилия, т. е. редко (табл. 4). Луга всегда граничат с лесом, поэтому единично встречаются виды из других биотопов, т. к. при длительном гниении крупной падали они активно передвигаются в поисках субстрата и находят приманку. Так, лесные виды навозников сем. Geotrupidae встречаются в лесу и на лугах (см. табл. 1).
Лесные биоценозы по сравнению с луговыми характеризуются более выровненным количественным составом жесткокрылых-некробионтов: индекс Пиелу Е здесь выше. Это обусловлено большей долей обычных и многочисленных видов.
Таблица 3. Количественные данные видового богатства жесткокрылых-некробионтов в биотопах южной Карелии
Table 3. Quantitative data of Coleoptera species collected from animal carcasses in different habitats of southern Karelia
| Индексы | Березняк | Сосняк | Луг |
| Всего видов | 53 | 49 | 58 |
| Индекс видового разнообразия Шеннона (He) | 3.42 | 3.35 | 3.31 |
| Индекс выравненности Пиелу (E) | 0.86 | 0.86 | 0.81 |
| Индекс доминирования Бергера – Паркера (d) | 0.09 | 0.08 | 0.1 |
| Число доминирующих видов (sH) | 10.63 | 10.21 | 9.92 |
Сопоставление фаунистических показателей трех биотопов показывает, что из 85 видов жесткокрылых-некробионтов общими оказываются всего 30 (35 %) (рис. 1). Сходство обеспечено обычными (2-й балл обилия) и многочисленными (3-й балл обилия) видами (см. табл. 1). Наибольшее качественное сходство имеет фауна луга с березняком разнотравно-черничным: для них отмечено 39 общих видов (68 %). Близость лесных биотопов подчеркивает и расчет евклидова расстояния, учитывающего обилие видов: большее сходство наблюдается между березняком и сосняком – dб/с = 8.77, а меньшее – этих биотопов с лугом: 8.89 и 8.83. То же показывает и корреляционный анализ: rб/с = 0.59, rл/с = 0.56, rб/л = 0.53, хотя различия коэффициентов незначимы.
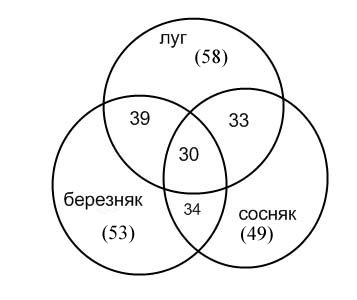
Рис. 1. Сходство жесткокрылых-некробионтов между биотопами по числу общих видов. В скобках количество видов, отмеченных в данном биотопе
Fig. 1. The resemblance of the biotopes of beetles collected from animals carcasses in the number of similar species. In brackets the number of species identified in every biotope is given
Таблица 4. Оценка встречаемости и относительного обилия видов жесткокрылых-некробионтов
Table 4. Evaluation of the occurrence and relative abundance of species of beetles collected from animal carcasses
| Балл, а | Вид по обилию | Количество видов, попадающих в диапазон интервала | ||
| В сосняке | В березняке | На лугу | ||
| 1 | Редкий (единичный) | 17 | 20 | 27 |
| 2 | Обычный | 20 | 25 | 21 |
| 3 | Многочисленный | 6 | 5 | 6 |
| 4 | Массовый | 5 | 3 | 4 |
Еще более рельефно визуализируются межбиотопические особенности фауны с помощью метода главных компонент (рис. 2).
Первая компонента регистрирует различие обилия животных на приманках разного размера. Так, например, средняя численность для M1 составляет 4.03 (максимальное удаление вправо), для B2 – 1.87 (максимальное удаление влево). Поскольку вариация размеров приманок по биотопам была практически одинакова, первая компонента (как и третья) почти совсем не различает сравниваемые списки.
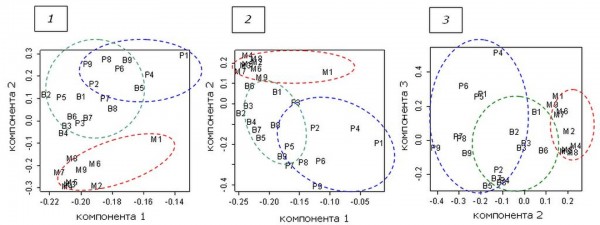
Рис. 2. Ординация фаунистических списков в осях факторных нагрузок. 1 – по всем данным (85 видов, абсолютные оценки), 2, 3 – без видов с очень высокой и нулевой численностью (29 видов, нормированные оценки обилия). Одна метка соответствует одной трупной приманке, расположенной в сосняке (P), березняке (B), на лугу (M)
Fig. 2. Ordination of faunal lists on the axes of the factor loadings. 1 – all data (85 species, the absolute value), 2, 3 – without species with a very high and zero abundance (29 species, normalized abundance estimates). One mark corresponds to one carcass bait located in a pine wood (P), birch wood (B), meadow (M)
По второй компоненте наблюдается хорошее разделение видовых списков. Луг занимает совершенно обособленное положение и не перекрывается с сосняком. Березняк занимает промежуточное положение, частично перекрывается и с сосняком, и с лугом. Нормирование (автошкалирование) почти не изменило картину, хотя сделало более определенной дифференциацию березняка и сосняка: степень трансгрессии снизилась – из области перекрытия списки B5 и B9 подтянулись ближе к центру своей группы.
Доказательство значимости наблюдаемых различий выполнялось с помощью бутстреп-анализа. Использована усеченная таблица нормированных данных по видам с невысоким и ненизким обилием (27 приманок, 29 видов). Всего выполнено три сравнения между тремя биотопами. В каждом случае различия по составу и обилию некрофагов оказались значимыми (табл. 5). Локальные фауны жуков-мертвоедов сосняка, березняка и луга различны.
Таблица 5. Сравнение редуцированных нормированных видовых списков некрофагов сосняка (P), березняка (B) и луга (M) методом бутстрепа (3000 циклов)
Table 5. Comparison of normalized reduced species lists of necrophages in a pine wood (P), birch wood (B) and meadow (M) by bootstrap method (3000 cycles)
| Расстояние | |||||
| Биотопы | расчетное | мин. | квантиль 2.5 % | квантиль 97.5 % | макс. |
| Сосняк / березняк | 4.388 | 1.000 | 1.582 | 3.280 | 4.388 |
| Сосняк / луг | 5.059 | 1.000 | 1.566 | 3.468 | 4.944 |
| Березняк / луг | 3.949 | 1.000 | 1.324 | 2.948 | 3.789 |
Обсуждение
Наблюдаемые различия имеют, на наш взгляд, вполне ясное экологическое объяснение.
Луговые биоценозы отличаются биотопической приуроченностью некоторых видов. Так, падальники р. Thanatophilus являются обитателями исключительно открытых ландшафтов. Виды р. Silpha в наших исследованиях встречались на лугах и в экотопах. Напротив, на Урале сильф отмечают в лесных биоценозах (Козьминых, Есюнин, 1990). На лугах также чаще отлавливается могильщик рыжебулавый Nicrophorus vespillo. Однако среди других двух видов жуков-могильщиков численность его низкая. Редко этот вид встречается и в Финляндии (Otronen, 1988). Напротив, в средней полосе N. vespillo довольно обычен в лесу (Трофимов, 2008).
Лесные биоценозы характеризуются высокой численностью трупоеда черного Necrodes littoralis, поверхностно-падального мертвоеда Oiceoptoma thoracicum, сапрафагов, обитающих в лесной подстилке, Deliphrum tectum, Tachinus marginellus. В сосняке в сухие годы на крупных приманках встречаются хищник Gnathoncus nanus и кожеед Dermestes ater, которые не отмечены в других биотопах.
Общими видами между биотопами являются жесткокрылые-сапрофаги Atheta paracrassicornis, A. сrassicornis, хищник Ontholestes murinus, паразитоид Aleochara curtula, кератофаг Necrobia violacea и др.
Заключение
В южной Карелии отмечено 85 видов жесткокрылых-некробионтов, из которых 35 % (30 видов) встречаются в любом биотопе. Наибольшее число видов выявлено во влажноразнотравном лугу – 58, здесь же обнаружено и наибольшее число оригинальных видов (12).
Своеобразные экологические условия в каждом биотопе формируют определенный комплекс жесткокрылых-некробионтов, различающийся видовым составом, численностью и соотношением таксономических групп.
Библиография
Волков А. Д. Типы леса Карелии. Петрозаводск: КарНЦ РАН, 2008. 180 с.
Козьминых В. О., Есюнин С. Л. Жуки-мертвоеды рода Silpha L. (Coleoptera, Silphidae) фауны Урала // Фауна и экология насекомых Урала: Межвуз. сб. науч. тр. Пермь, 1990. С. 94–105.
Коросов А. В. Экологические приложения компонентного анализа. Петрозаводск: Изд-во ПетрГУ, 1996. 152 с.
Коросов А. В. Специальные методы биометрии. Петрозаводск, 2007. 364 с.
Лябзина С. Н. Видовой состав и структура комплекса членистоногих-некробионтов Южной Карелии // Ученые записки Петрозаводского государственного университета. Сер.: Естественные и технические науки. 2011. № 4 (117). С. 10–19.
Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 184 с.
Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 286 с.
Раменская М. Л. Луговая растительность Карелии. Петрозаводск: Госиздат КАССР, 1958. 400 с.
Трофимов И. Е. Некоторые результаты популяционно-фенотипического анализа Nicrophorus vespillo (Coleoptera, Silphidae) из Калужского городского бора // Зоологический журнал. 2008. Т. 87. № 6. С. 658–664.
Шитиков В. К. Использование рандомизации и бутстрепа при обработке результатов экологических наблюдений // Принципы экологии. 2012. № 1. С. 4–24.
Kočárek P. Decomposition and Coleoptera succession on exposed carrion of small mammal in Opava, the Czech Republic // Eur. J. Soil. Biol. 2003. Vol. 39. P. 31-45.
Otronen M. The effect of body size on the outcome of fights in burying beetles (Nicrophorus) // Ann. Zool. Fen. 1988. Vol. 25. № 2. P. 191–201.
Благодарности
Авторы выражают благодарность В. Б. Семенову (Институт медицинской паразитологии и тропической медицины им. Е. И. Марциновского, ПМГМУ им. И. М. Сеченова г. Москва) за проверку и определение жуков сем. Staphylinidae.

 © 2011 - 2026
© 2011 - 2026